O papel da tríade na contração muscular
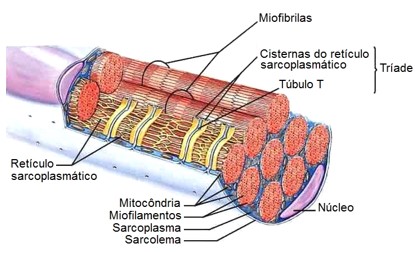
Em cada junção entre as bandas A e I, o sarcolema da fibra muscular estriada penetra a célula, formando um tubo alongado chamado de túbulo T (ou túbulo transversal).
Nos músculos esqueléticos, como cada túbulo T se projeta para para o interior da célula e se estende entre duas cisternas terminais do retículo sarcoplasmático (RS), forma as tríades, que permitem que a despolarização da membrana penetre rapidamente no interior da célula.
Os túbulos T passam de uma miofibrila à outra, circundando cada sarcômero. Assim, o sistema de túbulos T é responsável pela contração uniforme de cada fibra muscular esquelética.

A contração muscular depende da disponibilidade de íons Ca2+ citosólico, e o músculo relaxa quando o teor desse íon se reduz no sarcoplasma. Nas células musculares esqueléticas, o RS armazena e regula o fluxo de íons Ca2+.
Quando a membrana do RS é despolarizada pelo estímulo nervoso, os canais de Ca2+ se abrem, e esses íons, que estavam depositados nas cisternas do retículo, difundem-se passivamente (sem gasto de energia), atuando na troponina e possibilitando a interação da actina com a miosina. Quando cessa a despolarização, a membrana do RS, por processo ativo (que consome energia) e dependente de uma proteína de membrana Ca2+-ATPase, bombeia Ca2+ para o interior das cisternas do RS, o que interrompe a atividade contrátil e restabelece um reservatório de Ca2+ no retículo.
Inervação do músculo esquelético
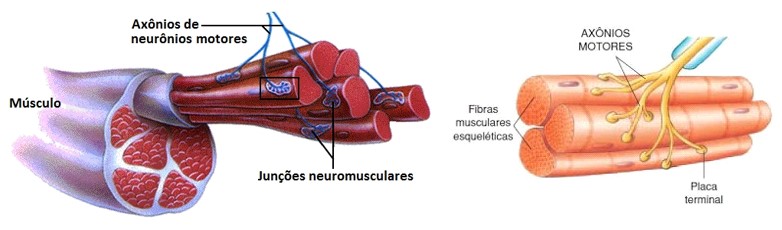
A contração muscular é essencialmente controlada por impulsos elétricos iniciados pelos nervos motores do sistema nervoso periférico somático, que trafegam ao longo do sarcolema. As fibras nervosas motoras que conduzem os impulsos do sistema nervoso central aos músculos esqueléticos se ramificam no tecido conjuntivo do perimísio, onde cada nervo origina numerosos ramos. No local de contato com a fibra muscular, o ramo final do axônio do nervo motor – terminal axonal ou botão terminal –, perde sua bainha de mielina e forma uma dilatação que se coloca dentro de uma depressão da superfície da fibra muscular. Essa estrutura chama-se placa motora ou junção neuromuscular (ou ainda junção mioneural).
OBS.:
Uma junção neuromuscular (ou sinapse neuromuscular) é uma região de sinapse química entre a parte terminal de um axônio motor (terminal axonal ou botão terminal) com uma placa motora, que é a região da membrana plasmática de uma fibra muscular (o sarcolema) onde se dá o contato entre o nervo motor e o músculo, permitindo desencadear a contração muscular.
O terminal axonal apresenta numerosas mitocôndrias e vesículas sinápticas com o neurotransmissor acetilcolina (ACh).
Na junção neuromuscular, o sarcolema forma as dobras juncionais (apresenta-se pregueado). Quando uma fibra do nervo motor recebe um impulso nervoso, o terminal axonal libera acetilcolina, que se difunde através da fenda sináptica e se liga aos receptores situados no sarcolema das dobras juncionais. A ligação dos receptores com o neurotransmissor faz com que o sarcolema torne-se mais permeável ao sódio, o que resulta na sua despolarização.
A despolarização iniciada na placa motora propaga-se ao longo da membrana da fibra muscular e penetra o interior da fibra através do sistema de túbulos T.
 |  |
Em cada tríade o sinal despolarizador passa para o retículo sarcoplasmático, resultando na liberação de Ca2+, que inicia o ciclo de contração. Quando a despolarização termina, o Ca2+ é transportado ativamente de volta para as cisternas do retículo sarcoplasmático, e a fibra muscular relaxa.
OBS.:
O excesso de acetilcolina é hidrolisado pela colinesterase encontrada na fenda sináptica. A destruição da acetilcolina é necessária para evitar o contato prolongado do neurotransmissor com os receptores do sarcolema.
Acoplamento Excitação-Contração
O termo acoplamento excitação-contração se refere ao mecanismo pelo qual o potencial de ação provoca a contração das miofibrilas, ou seja, como o sinal elétrico (potencial de ação) é convertido em contração muscular. Assim, o termo designa todo o processo que ocorre desde a despolarização da membrana da fibra muscular até a liberação de íons cálcio dos estoques intracelulares do RS, que culmina no encurtamento do músculo via atividade dos miofilamentos.
Para facilitar o entendimento, abordaremos o acoplamento excitação-contração em duas etapas:
- mecanismo geral de contração muscular
- mecanismo molecular da contração muscular
Mecanismo geral de contração muscular
O início e a execução de uma contração muscular ocorre nas seguintes etapas sequenciais:
1- Os potenciais de ação cursam pelo nervo motor até suas terminações nas fibras musculares.
 | 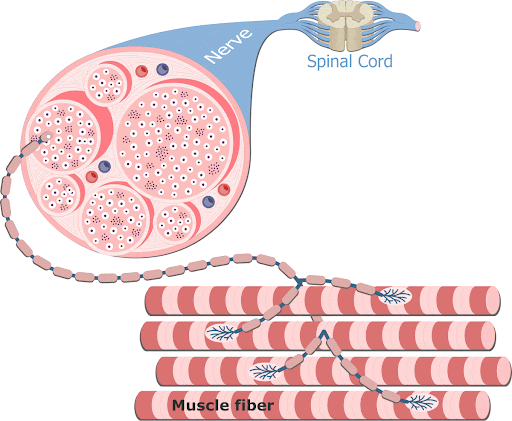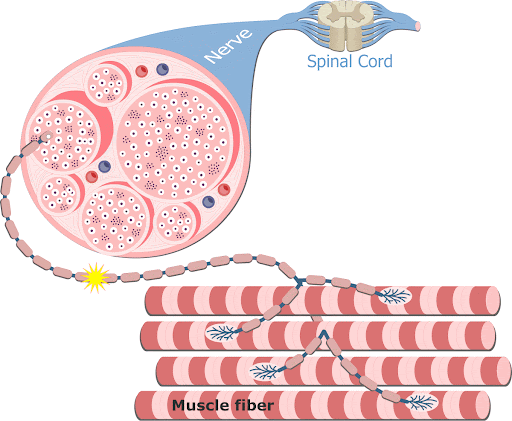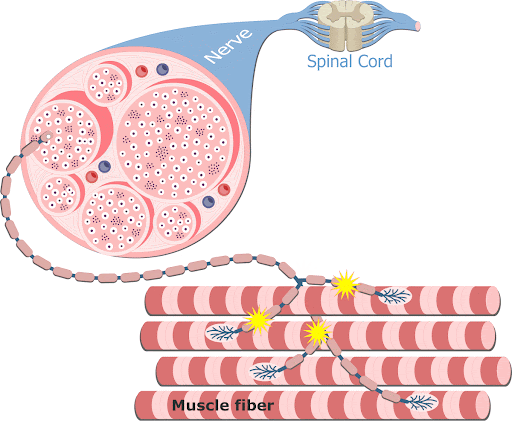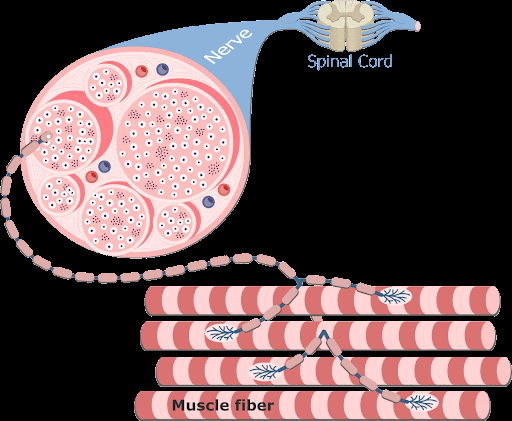 |
2- Em cada terminação, o impulso nervoso desencadeia liberação de acetilcolina do botão sináptico (expansão do terminal axonal) na fenda sináptica.
3- A acetilcolina se liga a receptores específicos na placa motora da junção neuromuscular, abrindo canais de sódio regulados por acetilcolina.
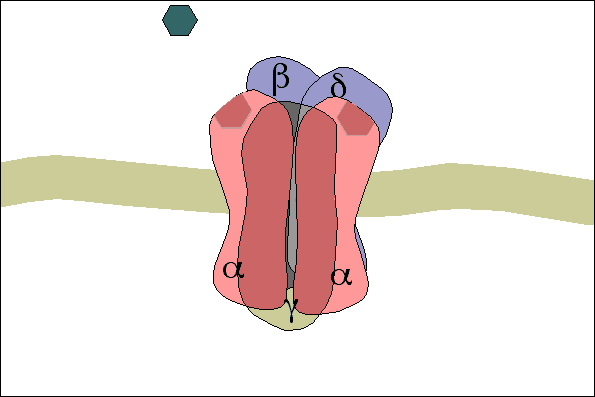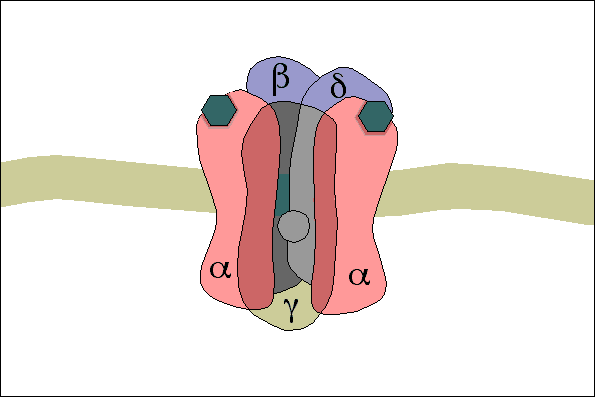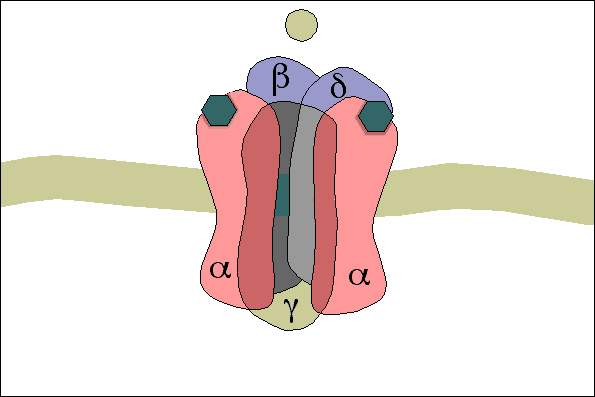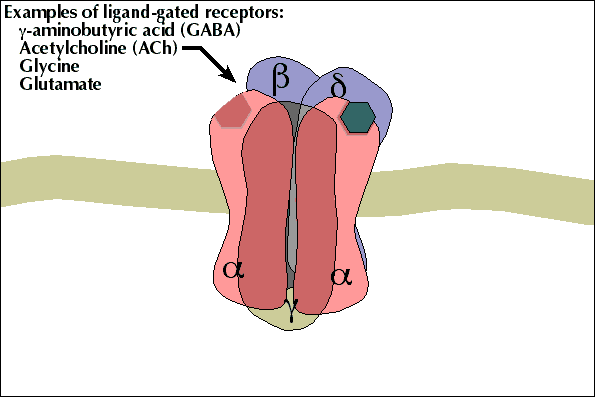
4- A abertura desses canais permite a difusão de grande quantidade de íons sódio para o lado interno da membrana das fibras musculares. Isso desencadeia o potencial de ação na membrana.
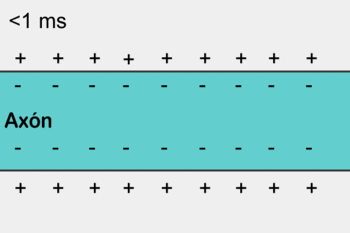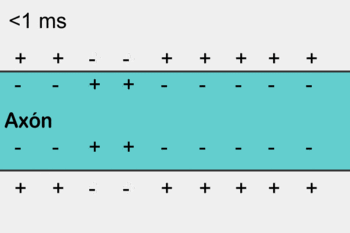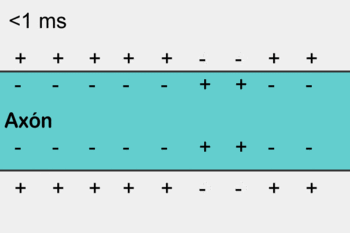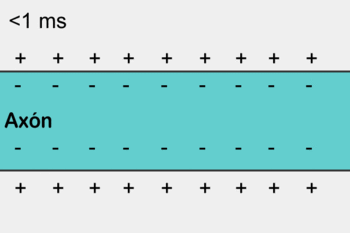
5- O potencial de ação se propaga por toda a membrana da fibra muscular do mesmo modo como o potencial de ação cursa pela membrana das fibras nervosas.
 | 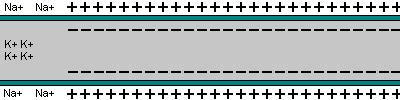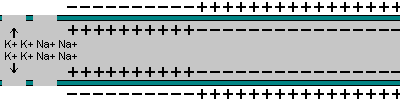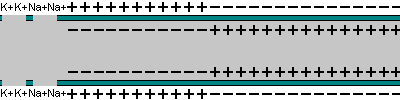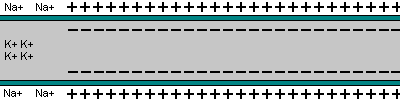 |
6- O potencial de ação despolariza a membrana muscular e se propaga pelos túbulos T, fazendo com que o retículo sarcoplasmático libere grande quantidade de íons cálcio para o sarcoplasma.
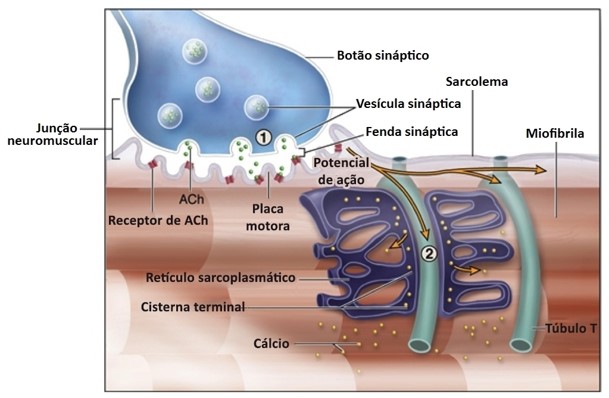
7- Os íons cálcio ativam as forças atrativas entre os filamentos de miosina e actina, fazendo com que eles deslizem ao lado um do outro, que é o processo contrátil.
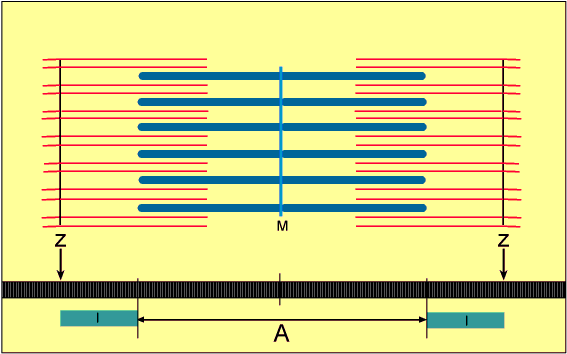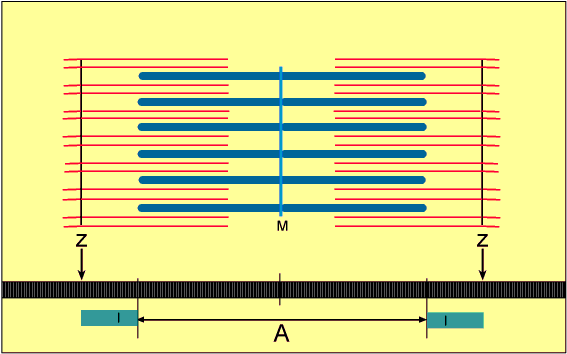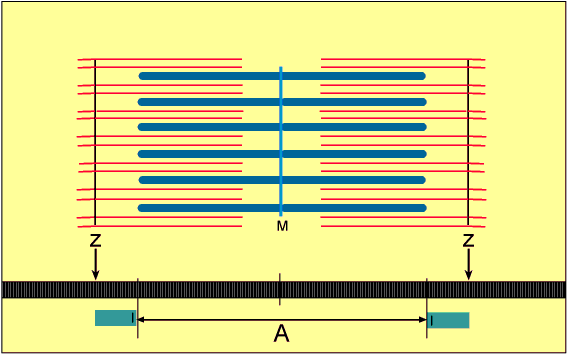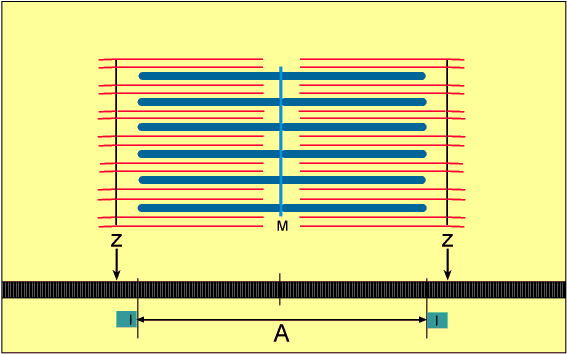
8- Assim que cessa o estímulo, os íons cálcio são bombeados de volta para dentro do retículo sarcoplasmático pela bomba de cálcio presente na sua membrana, permanecendo aí armazenados até que novo potencial de ação muscular se inicie. Essa retirada de íons cálcio das miofibrilas faz com que a contração muscular cesse.
Mecanismo molecular da contração muscular
No estado relaxado, o sarcômero consiste em filamentos finos e grossos que se sobrepõem parcialmente. Também, as extremidades dos filamentos de actina que se estendem de duas linhas Z sucessivas mal se sobrepõem.
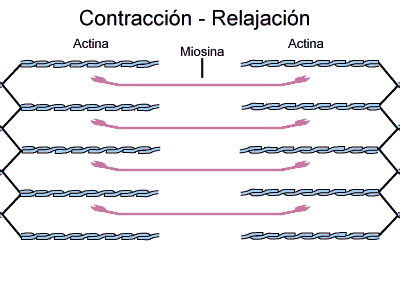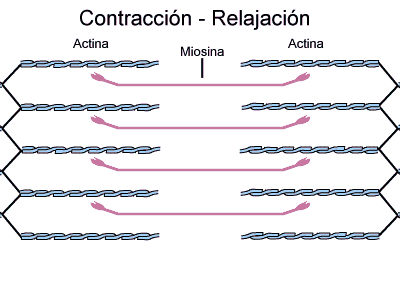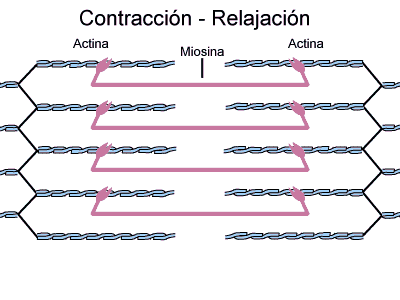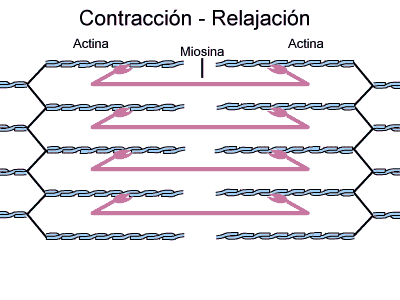
Inversamente, no estado contraído, apesar de os filamentos finos e grossos conservarem seus comprimentos originais, os filamentos de actina são tracionados por entre os filamentos de miosina, de forma que suas extremidades se sobrepõem umas às outras em sua extensão máxima. Também as linhas Z são tracionadas pelos filamentos de actina até as extremidades dos filamentos de miosina. Desse modo, a contração muscular ocorre por mecanismo de deslizamento dos miofilamentos uns sobre os outros, o que aumenta o tamanho da zona de sobreposição entre os filamentos e diminui o tamanho do sarcômero.

Isto resulta da força gerada pela interação das pontes cruzadas dos filamentos de miosina com os filamentos de actina.
Mecanismo da contração
A contração se inicia na banda A, onde os filamentos finos e grossos se sobrepõem.
Durante o ciclo de contração a actina e a miosina interagem da seguinte maneira:
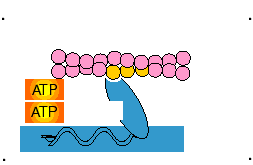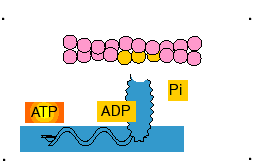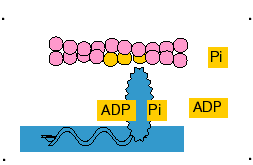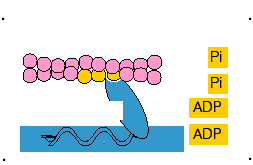
1- Durante o repouso, o ATP liga-se à cabeça da miosina na região de ligação ao ATP, que é dotada de atividade ATPásica. Porém, para clivar a molécula de ATP e liberar energia, a miosina necessita da actina, que atua como cofator. Nesta etapa, a conformação da cabeça é tal que se estende em direção ao filamento de actina, mas ainda está ligada à actina.
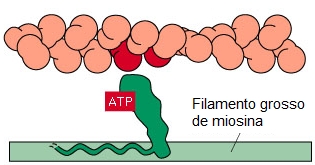
2- No músculo em repouso a miosina não pode associar-se à actina devido à repressão do local de ligação pelo complexo troponina-tropomiosina fixado sobre o filamento de actina, mais especificamente, pela subunidade TnI da troponina que cobre a região de ligação da actina com a miosina, onde ocorre a interação dos filamentos.
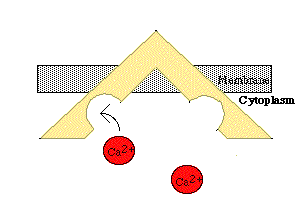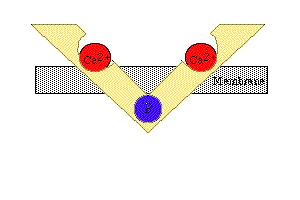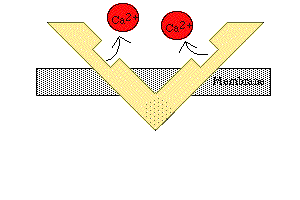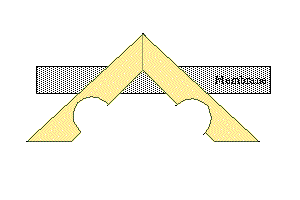
3- Quando há disponibilidade de íons Ca2+, esses se combinam com a unidade TnC da troponina, mudando a configuração espacial das três subunidades de troponina e empurrando a molécula de tropomiosina mais para dentro do sulco da hélice de actina. Isto expõe a região de ligação da actina com a miosina, ocorrendo, então, a interação entre os filamentos.
4- Como resultado, a ATPase das cabeças da miosina é ativada, clivando o ATP em ADP e fosfato inorgânico (Pi) e liberando energia (o ADP permanece ligado à cabeça). Isto promove uma deformação da cabeça e de parte do bastão da miosina nas pontes cruzadas, aumentando a curvatura da cabeça. Como a actina está combinada com a miosina, o movimento da cabeça da miosina empurra o filamento da actina, promovendo seu deslizamento sobre o filamento de miosina.
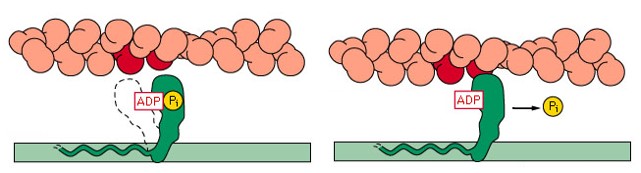
5- Uma vez em que a cabeça da miosina esteja inclinada, isso permite a liberação do ADP que estava ligado à cabeça.
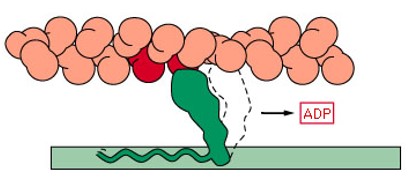
6- No local onde foi liberado o ADP, nova molécula de ATP se liga. Esta ação determina a volta da cabeça de miosina para sua posição primitiva e seu consequente desligamento da cabeça da actina, preparando-se para novo ciclo.