
Fisiologia da respiração
Ventilação pulmonar
Os pulmões estão posicionados dentro da cavidade torácica, a qual é definida pelo gradil costal e pela coluna vertebral. Os pulmões são rodeados por vários músculos que contribuem para a respiração.
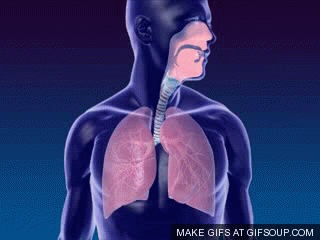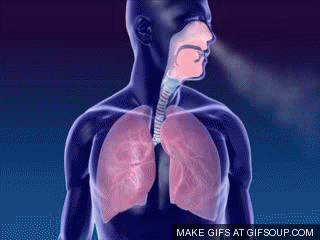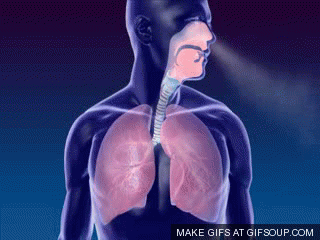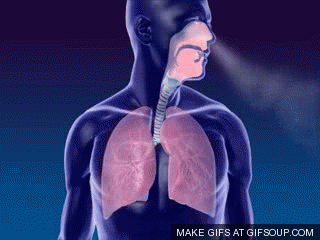
A primeira troca na fisiologia respiratória é a ventilação ou respiração, ou seja, o fluxo de ar entre a atmosfera e os alvéolos (ou a entrada e saída de ar dos pulmões). Um único ciclo respiratório consiste em uma inspiração seguida por uma expiração. Assim, respiração ou ventilação pulmonar é composta por duas fases:
- inspiração, o período no qual o ar flui para dentro dos pulmões,
- expiração, o período no qual o ar sai dos pulmões.
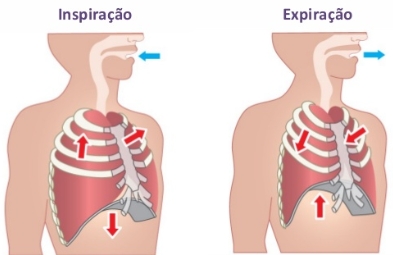
A inspiração, que promove a entrada de ar nos pulmões, dá-se pela contração da musculatura do diafragma e dos músculos intercostais. O diafragma abaixa e as costelas elevam-se, promovendo o aumento da caixa torácica, com consequente redução da pressão interna (em relação à externa), forçando o ar a entrar nos pulmões.
A expiração, que promove a saída de ar dos pulmões, dá-se pelo relaxamento da musculatura do diafragma e dos músculos intercostais. O diafragma eleva-se e as costelas abaixam, o que diminui o volume da caixa torácica, com consequente aumento da pressão interna, forçando o ar a sair dos pulmões.
Lembrando que a ventilação pulmonar é um processo mecânico que depende de modificações do volume da cavidade torácica.
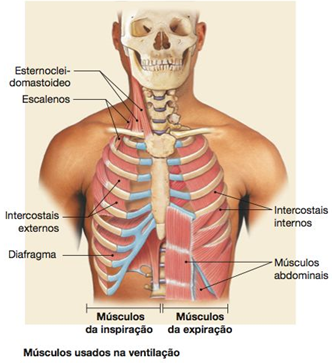
Os músculos da respiração e a parede torácica são componentes essenciais do sistema respiratório. Os pulmões não são capazes de insuflarem a si próprios – a força para essa insuflação deve ser fornecida pelos músculos respiratórios. Todos os músculos que elevam a caixa torácica são classificados como músculos da inspiração, e aqueles que deprimem a caixa torácica são classificados como músculos da expiração. A parede torácica deve estar intacta e ser capaz de expandir para o ar entrar nos alvéolos normalmente.
Os pulmões podem ser expandidos e contraídos de duas maneiras:
- por movimentos de subida e descida do diafragma para aumentar ou diminuir a cavidade torácica,
- pela elevação e depressão das costelas para aumentar e diminuir o diâmetro anteroposterior da cavidade torácica.
As interações entre os músculos respiratórios e a parede torácica na ventilação pulmonar são mostradas ao abaixo.
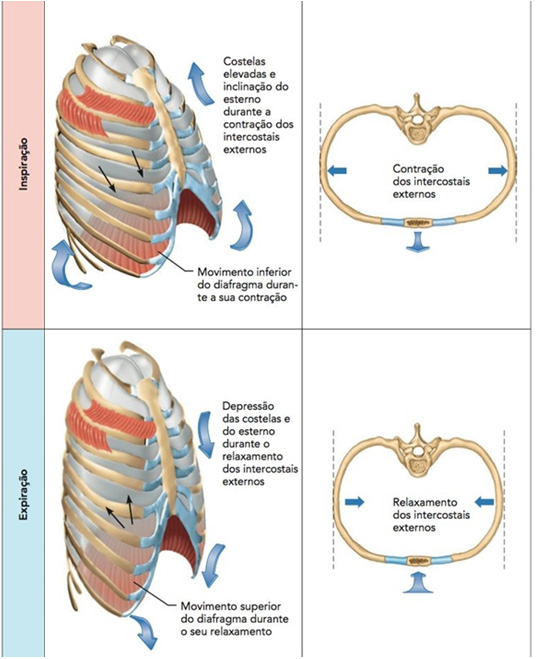
A respiração tranquila normal é realizada quase inteiramente pelo movimento do diafragma. Durante a inspiração, a contração diafragmática puxa as superfícies inferiores dos pulmões para baixo. Depois, durante a expiração, o diafragma simplesmente relaxa, e o recuo elástico dos pulmões, parede torácica e estruturas abdominais comprime os pulmões e expele o ar.
Resumindo:
- inspiração: contração do diafragma e dos músculos intercostais externos diafragma abaixa e costelas elevam-se aumento da caixa torácica redução da pressão interna entrada do ar.
- expiração: relaxamento do diafragma e dos músculos intercostais externos diafragma eleva-se e costelas abaixam diminui o volume da caixa torácica aumento da pressão interna saída do ar.
Transporte de gases respiratórios
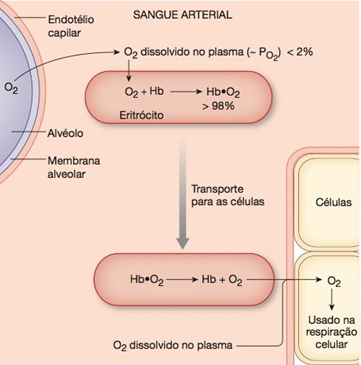
O transporte de gás oxigênio está a cargo da hemoglobina, proteína presente nas hemácias. Cada molécula de hemoglobina combina-se com 4 moléculas de gás oxigênio, formando a oxiemoglobina.
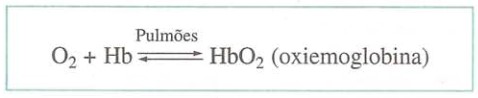
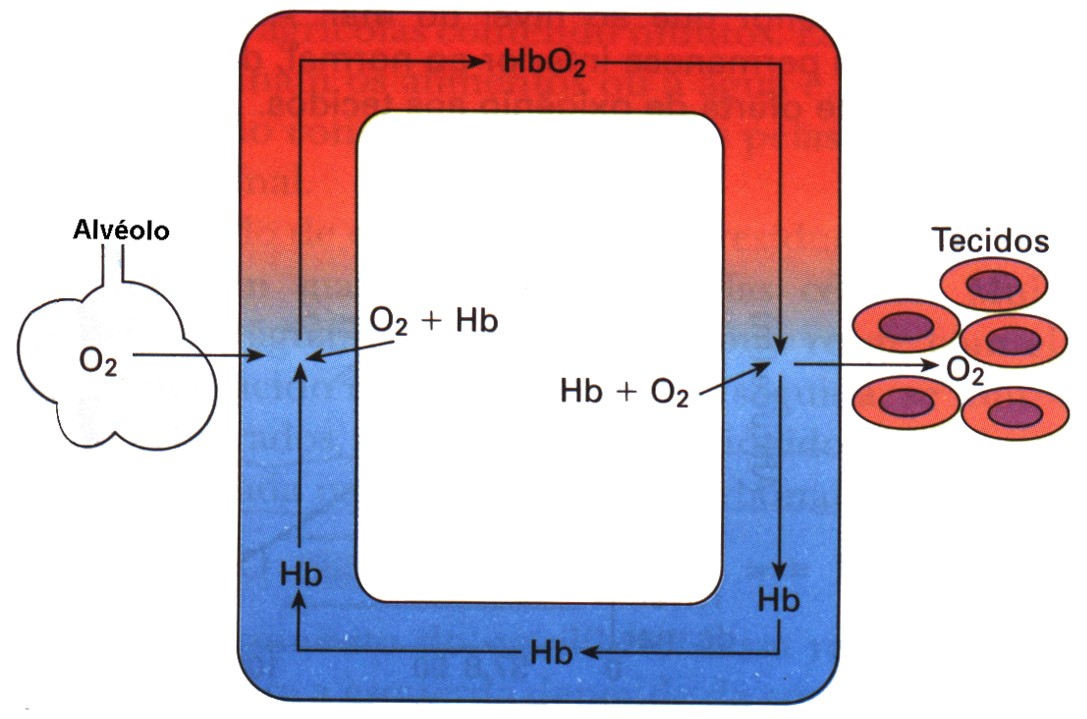
Nos alvéolos pulmonares o gás oxigênio do ar difunde-se para os capilares sanguíneos e penetra nas hemácias, onde se combina com a hemoglobina, enquanto o gás carbônico (CO2) é liberado para o ar (processo chamado hematose). Nos tecidos ocorre um processo inverso: o gás oxigênio dissocia-se da hemoglobina e difunde-se pelo líquido tissular, atingindo as células.
O dióxido de carbono (gás carbônico) é um subproduto da respiração celular. Assim, o transporte de gases no sangue inclui a remoção de dióxido de carbono das células, bem como a entrega de oxigênio para as células.
O CO2 difunde-se dos tecidos para o sangue, e parte dele se une à hemoglobina livre, formando um composto também instável denominado carboemoglobina.

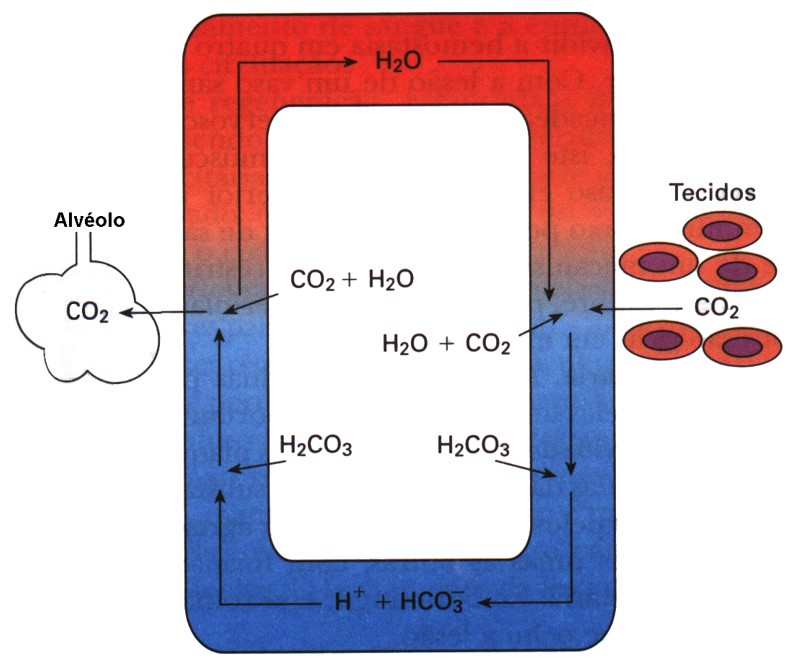
Nos pulmões, CO2 difunde-se para os alvéolos, deixando a hemoglobina livre. Enquanto praticamente todo o O2 é transportado pela hemoglobina, isto não ocorre com o CO2. As células produzem muito mais CO2 do que pode dissolver-se no plasma. Assim, o sangue transporta o CO2 dos tecidos para os pulmões sob três formas:
(1) apenas cerca de 7-10% do CO2 transportado pelo sangue venoso são dissolvidos no plasma;
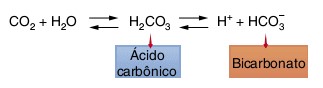
(2) os 90-93% restantes difundem-se para os eritrócitos (hemácias), onde aproximadamente 70% são convertidos no íon bicarbonato (HCO3‒). Devido à presença da anidrase carbônica. O H2CO3 é instável e rapidamente se dissocia em íons hidrogênio (H+) e íons bicarbonato (HCO3−).
(3) pouco mais de 20% (~23%) ligam-se à hemoglobina (Hb-CO2). Nesta forma, o CO2 é carregado nos eritrócitos como carboemoglobina.
OBS.:
Quando o CO2 se difunde para os eritrócitos, ele se combina com a água, formando ácido carbônico (H2CO3), devido à presença da anidrase carbônica. O H2CO3 é instável e rapidamente se dissocia em íons hidrogênio (H+) e íons bicarbonato (HCO3−).
Apesar desta reação também acontecer no plasma, ela é milhares de vezes mais rápida nos eritrócitos pela presença da anidrase carbônica, uma enzima que reversivelmente catalisa a conversão do dióxido de carbono e água a ácido carbônico.
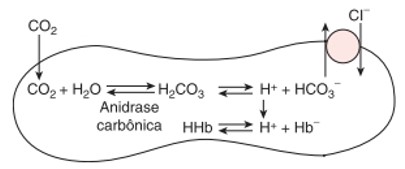
O H+ é tamponado (neutralizado), principalmente pela hemoglobina, enquanto o HCO3− difunde-se rapidamente para o plasma.
Os íons bicarbonato transportados livres no plasma contribuem para que o pH do sangue arterial e venoso seja ligeiramente básico. Entretanto, a pequena quantidade de ácido carbônico formada no plasma, em função da união do gás carbônico com a água, é responsável por ligeira diminuição do pH do sangue venoso, que passa a ser 7,36, enquanto o sangue arterial tem pH 7,4. Nos dois casos, entretanto, o pH se mantém ligeiramente básico.
Nos pulmões, ocorre o processo inverso. Assim, o bicarbonato que está no plasma penetra novamente os eritrócitos (e os cloretos voltam para o plasma), se liga com o H+ para formar ácido carbônico, que em seguida é clivado pela anidrase carbônica para liberar CO2 e água. Este CO2, juntamente com o liberado da hemoglobina e o dissolvido no plasma, se difunde do sangue para os alvéolos, a favor de seu gradiente de concentração (pressão parcial).
Saturação da hemoglobina
A combinação da hemoglobina (Hb) com o oxigênio, chamada de oxiemoglobina, é escrita HbO2. A hemoglobina libera o oxigênio e passa a se chamar hemoglobina reduzida ou desoxiemoglobina, e é escrita HHb. A associação e a dissociação do O2 podem ser indicadas por uma única equação reversível.

Após a primeira ligação de uma molécula de O2 com o ferro, a molécula de hemoglobina (Hb) modifica sua forma. Como resultado, a Hb rapidamente liga mais duas moléculas, e a ligação com a quarta molécula é ainda mais facilitada. Quando todos os grupos heme estiverem ligados aos O2, considera-se que a molécula de hemoglobina está totalmente saturada. Quando uma, duas ou três moléculas de oxigênio estão ligadas, a hemoglobina está parcialmente saturada. Seguindo o mesmo raciocínio, a liberação de uma molécula de oxigênio facilita a liberação da próxima e, assim, sucessivamente. Assim, a afinidade (força da ligação) da hemoglobina com o oxigênio é modificada pelo grau da saturação de oxigênio e, consequentemente, tanto a associação quanto a dissociação do oxigênio ocorre de forma muito eficiente.
Curva de dissociação da hemoglobina (Hb)
A proporção de oxigênio ligado à hemoglobina é expressa como percentual de saturação. Esse percentual corresponde ao conteúdo de oxigênio ligado à hemoglobina dividido pela capacidade de oxigênio da hemoglobina no sangue multiplicado por 100%.
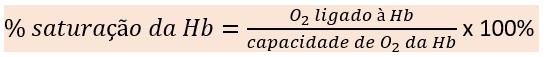
A capacidade de oxigênio de um indivíduo depende da quantidade de hemoglobina no sangue. O conteúdo de oxigênio no sangue também depende da quantidade de hemoglobina presente, bem como da pressão parcial desse gás (PO2). Tanto o conteúdo como a capacidade de oxigênio são expressos em mililitros de oxigênio por 100 mL de sangue. Por outro lado, o percentual de saturação da hemoglobina expressa apenas um percentual, e não uma quantidade ou volume de oxigênio, ou seja, o “percentual de saturação” não é o mesmo que o “conteúdo de oxigênio”. Por exemplo, dois pacientes podem ter o mesmo percentual de saturação da hemoglobina, mas, se um deles tiver uma menor concentração de hemoglobina no sangue devido a uma anemia, ele terá um menor conteúdo de oxigênio no sangue.
A relação entre a PO2 do plasma e o percentual de saturação da hemoglobina pode ser expressa graficamente como a curva de dissociação da oxiemoglobina. Como a afinidade da hemoglobina pelo O2 é modificada com a ligação do O2, a relação entre o grau de saturação de hemoglobina e a PO2 do sangue não é linear, como mostra a figura da curva de dissociação de oxigênio-hemoglobina. Esta curva apresenta um formato sigmoide e tem uma inclinação ascendente nos valores de PO2 entre 10 e 50 mmHg e, então, atinge um platô entre 70 e 100 mmHg. Observe que a curva é quase plana em níveis de PO2 maiores que 100 mmHg (isto é, a inclinação se aproxima de zero). A uma PO2 acima de 100 mmHg, mesmo grandes mudanças na PO2 causam apenas mudanças pequenas no percentual de saturação. De fato, a hemoglobina não é 100% saturada até a PO2 alcançar aproximadamente 650 mmHg, uma pressão parcial muito mais alta do que encontramos na vida cotidiana.

Intoxicação por monóxido de carbono (CO)

O monóxido de carbono, liberado pela “queima” incompleta de combustíveis fósseis e pela fumaça dos cigarros entre outros, combina-se com a hemoglobina de uma maneira mais estável do que o oxigênio, formando o carboxiemoglobina. Dessa forma, a hemoglobina fica impossibilitada de transportar o oxigênio, podendo levar à morte por asfixia. Veja as tabelas abaixo, retiradas da prova do ENEM de 98:
Um dos índices de qualidade do ar diz respeito à concentração de monóxido de carbono (CO), pois esse gás pode causar vários danos à saúde. A tabela abaixo mostra a relação entre a qualidade do ar e a concentração de CO.

Para analisar os efeitos do CO sobre os seres humanos, dispõe-se dos seguintes dados:
