
Tipos básicos de resposta imune
As respostas que desenvolvemos contra infecções por patógenos potenciais são conhecidas como respostas imunes. A defesa contra os microrganismos é mediada pelas reações iniciais da imunidade inata, responsável pela proteção inicial contra as infecções, e pelas respostas mais tardias da imunidade adquirida, que se desenvolve mais lentamente e é responsável pela defesa mais eficaz contra as infecções.
A imunidade inata (também chamada de imunidade natural ou nativa) consiste em mecanismos que existem antes da infecção e que são capazes de gerar respostas rápidas aos microrganismos, reagindo essencialmente do mesmo modo a qualquer tipo de infecção.
A expressão imunidade inata refere-se ao fato de que esse tipo de defesa está sempre presente nos indivíduos saudáveis, estando preparada para bloquear a entrada de microrganismos e eliminar rapidamente aqueles que conseguem entrar nos tecidos do hospedeiro. Desta forma, os mecanismos da imunidade inata não distinguem um agente infeccioso de outro: os componentes da imunidade inata reconhecem estruturas que são comuns a diversas classes de microrganismos relacionados (podem não distinguir diferenças discretas entre microrganismos) e que não estão presentes nas células do hospedeiro (aloantígenos).
A característica em comum dos mecanismos da imunidade inata é que eles reconhecem e respondem aos microrganismos, mas não reagem a substâncias não-bacterianas. Ela também pode ser desencadeada pelas células do hospedeiro que são danificadas pelos microrganismos.
Os componentes da imunidade inata formam o sistema imunológico inato. Os principais componentes da imunidade inata são:
- barreiras físicas e químicas, como os epitélios e as substâncias químicas antimicrobiana produzidas nas superfícies epiteliais;
- células fagocitárias, células dendríticas e células matadoras naturais (natural killer ‒ NK);
- proteínas do sangue, incluindo membros do sistema complemento e outros mediadores da inflamação;
- proteínas denominadas citocinas, que regulam e coordenam muitas das atividades das células da imunidade natural.
Os mecanismos da imunidade inata incluem duas linhas de defesa que os invasores encontram ao tentarem penetrar o corpo. A primeira linha, mais externa, é formada pela pele e pelas mucosas dos sistemas respiratório, digestório e urogenital (barreiras epiteliais, células especializadas e antibióticos naturais, que bloqueiam a entrada dos microrganismos). Se um microrganismo conseguir vencer essas barreiras, penetrar no epitélio e entrar nos tecidos ou na circulação, enfrentará a segunda linha de defesa inespecífica (interna): serão atacados pelos fagócitos, células natural killer (NK) e diversas proteínas plasmáticas, incluindo as proteínas do sistema do complemento, os quais agem imediatamente após a infecção.
Além de fornecer a defesa inicial contra as infecções, as respostas da imunidade inata estimulam as respostas da imunidade adquirida contra os agentes infecciosos. |
Apesar de a imunidade inata poder combater muitas infecções de maneira eficaz, microrganismos patogênicos (capazes de causar doença) evoluíram para resistir aos seus mecanismos. A defesa contra esses agentes infecciosos é função da resposta imunológica adquirida, e é por isso que defeitos nesse sistema resultam em maior suscetibilidade a infecções.
A imunidade adquirida (também chamada de imunidade adaptativa) é o tipo de defesa estimulada pelos microrganismos que invadem os tecidos, adaptando-se à presença dos invasores microbianos. Os mecanismos da imunidade adquirida constituem a terceira e última linha de defesa, em que as respostas não são indiscriminadas, mas específicas. Participam desses mecanismos os linfócitos T e B e seus produtos, como os anticorpos.
Como visto anteriormente, os mecanismos da imunidade inata reconhecem estruturas comuns a classes de microrganismos, enquanto as células da imunidade adquirida, ou seja, os linfócitos T e B, expressam receptores que reconhecem especificamente diversas substâncias produzidas pelos microrganismos, assim como moléculas não-infecciosas.
As respostas imunológicas adquiridas só são desencadeadas se os microrganismos ou seus antígenos passarem pelas barreiras epiteliais e forem transportados para os órgãos linfoides, onde podem ser reconhecidos pelos linfócitos. As respostas imunológicas adquiridas geram mecanismos especializados para o combate de diversos tipos de infecção. Por exemplo, os anticorpos eliminam os microrganismos presentes nos líquidos extracelulares (resposta imune humoral), enquanto os linfócitos T ativados eliminam os microrganismos que vivem dentro das células (resposta imune celular).

As respostas adquiridas geralmente usam células e moléculas do sistema imunológico inato para eliminar os microrganismos, e funções imunológicas adquiridas para aumentar acentuadamente esses mecanismos antimicrobianos da imunidade inata. Por exemplo, os anticorpos (um componente da imunidade adquirida) se ligam aos microrganismos que, quando revestidos pelos anticorpos, se ligam avidamente às células fagocitárias (um componente da imunidade inata) ativando-as e sendo fagocitados e destruídos por elas.
Características da imunidade inata e da adquirida | ||
| Características | Inata | Adquirida |
| Especificidade | Para estruturas compartilhadas por grupos de microrganismos relacionados | Para antígenos microbianos e não microbianos |
| Diversidade | Limitada | Muito grande |
| Memória | Nenhuma | Sim |
| Não reação ao próprio | Sim | Sim |
Componentes da imunidade inata e da adquirida | ||
| Componentes | Inata | Adquirida |
| Barreiras físicas e químicas | Pele, epitélio das mucosas, substâncias químicas antimicrobianas | Linfócitos nos epitélios, anticorpos secretados nas superfícies epiteliais |
| Proteínas sanguíneas | Complemento | Anticorpos (Imunoglobulinas) |
| Células | Fagócitos, células dendríticas, células NK | Linfócitos |
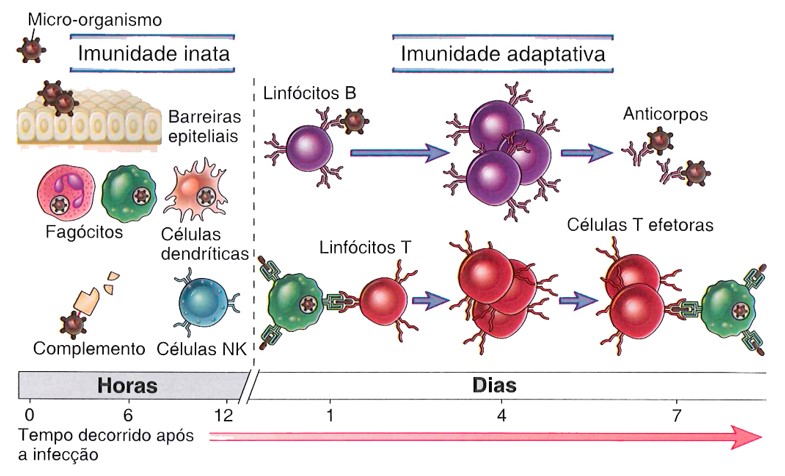
Além de fornecer a defesa inicial contra infecções, a imunidade inata também instrui o sistema imunológico adquirido a responder aos diversos microrganismos de maneira eficaz. Por sua vez, a resposta imunológica adquirida geralmente usa mecanismos da imunidade inata para erradicar infecções. Assim, existe uma comunicação bidirecional constante entre a imunidade inata e a imunidade adquirida. Por essas razões, há um grande interesse na definição dos mecanismos da imunidade inata para que se possa aprender a como aproveitá-los para otimizar a defesa contra as infecções.
Componentes e mecanismos da imunidade inata
O sistema imunológico inato consiste em epitélio, que fornece barreiras às infecções, células na circulação e nos tecidos e diversas proteínas plasmáticas. Assim, fagocitose, liberação de mediadores inflamatórios, ativação de proteínas do sistema complemento, bem como síntese de proteínas de fase aguda, citocinas e quimiocinas são os principais mecanismos na imunidade inata. Esses componentes desempenham papéis diversos, mas complementares, bloqueando a entrada dos microrganismos e eliminando aqueles que entram nos tecidos do hospedeiro.
Barreiras epiteliais: 1ª linha de defesa
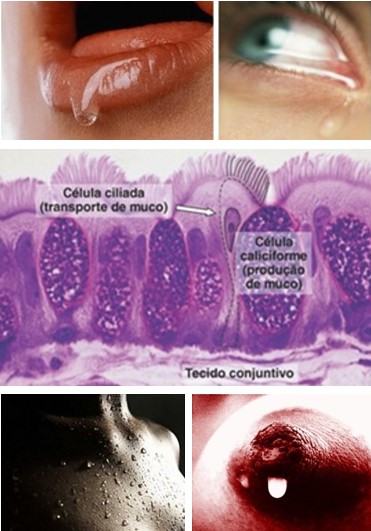
A pele, o trato gastrointestinal e o trato respiratório são as três principais interfaces entre o corpo e o ambiente externo e, consequentemente, as portas de entrada frequentes dos microrganismos do ambiente externo, pois esses podem entrar através dessas interfaces pelo contato físico, ingestão ou respiração. Desta forma, como previamente mencionado, o organismo possui barreiras naturais físicas e químicas que são obviamente inespecíficas, como a da pele (queratina, lipídios e ácidos graxos), a saliva, o ácido clorídrico do estômago, o pH da vagina, a cera da orelha externa, muco presente nas mucosas e no trato respiratório, cílios do epitélio respiratório, peristaltismo, microbiota normal, entre outros.
Todas as três portas de entrada são revestidas por um epitélio contínuo que interfere fisicamente na entrada dos patógenos. Além do tecido epitelial estratificado pavimentoso queratinizado, os ácidos graxos livres produzidos nas glândulas sebáceas e por microrganismos presentes na superfície da pele, o pH baixo (entre 5,5 e 6,0), a salinidade do suor e o ambiente relativamente seco da pele proporcionam condições desfavoráveis para a sobrevivência da maioria dos microrganismos.
As células epiteliais também produzem antibióticos peptídicos que destroem as bactérias. A lisozima, por exemplo, é uma enzima encontrada em vários fluidos corpóreos, como a saliva, as lágrimas, o muco brônquico e o suor, cuja atividade antimicrobiana é bem conhecida: atua catalisando a hidrólise de polissacarídeos da parede celular e causando a lise das bactérias devido às condições hiposmóticas da saliva ou pela ação de outros componentes salivares antimicrobianos. Um outro exemplo é a lactoferrina, segunda proteína predominante no leite humano e que apresenta atividades bacteriostática, bactericida, fungicida, antiviral, antiparasítica, anti-inflamatória e imunomoduladora.
As demais barreiras naturais citadas atuam das seguintes formas:
- o muco presente nas mucosas forma um manto pegajoso e lubrificante que age como uma barreira físico-química, auxiliando no aprisionamento de patógenos ou de suas substâncias, na inibição do crescimento ou na lise desses microrganismos;
- os cílios do epitélio respiratório criam uma corrente suave que move o muco contaminado interiormente para a parte oral da faringe, onde será deglutido e digerido pelo suco gástrico;
- a bile e ambiente ácido do estômago, da bexiga urinária e da vagina inativam muitos vírus e bactérias;
- a cera da orelha externa retém poeira e micróbios que eventualmente entram nas orelhas;
- o peristaltismo e fluxo urinário limitam o estabelecimento da infecção;
- a microbiota normal promove proteção e defesa do organismo contra a tentativa de colonização de patógenos e também a estimulação do sistema imune em recém-nascidos.
Inflamação: 2ª linha de defesa
Se as barreiras naturais do corpo forem vencidas (1ª linha de defesa), o combate ao agente infeccioso entra em outra fase (2ª linha de defesa), que envolve as células que participam da imunidade inata (mastócitos, leucócitos granulócitos, monócitos, macrófagos, células NK e células dendríticas), proteínas do sangue, incluindo membros do sistema complemento e outros mediadores da inflamação, como as citocinas, que regulam e coordenam muitas das atividades das células da imunidade natural.
.jpg?alt=media&token=b2943d2c-9839-4c44-9468-33d7c2765d78)
Os microrganismos que rompem as defesas das barreiras epiteliais encontram células e moléculas que imediatamente desenvolvem uma resposta imune inata. Os macrófagos que encontram bactérias ou outros patógenos nos tecidos são ativados para liberar citocinas que aumentam a permeabilidade dos vasos sanguíneos permitindo que fluidos e proteínas passem para os tecidos. Essas células ainda liberam quimiocinas, pequenas citocinas que direcionam a migração dos neutrófilos e de outras células imunes, tais como monócitos, basófilos, eosinófilos, células NK e linfócitos T, para o local de infecção. As quimiocinas também controlam o tráfego de células imunes inatas entre a medula óssea, o sangue e os tecidos periféricos durante a inflamação. Além disso, a adesão das células endoteliais dos vasos sanguíneos é alterada de modo que os fagócitos do sangue aderem à parede e são capazes de passar por ela (diapedese): primeiro os neutrófilos e depois os monócitos passam do sangue para os tecidos, como mostrado na figura abaixo.
.jpg?alt=media&token=ae3ee26c-0ce4-4d75-a5d9-c8e581dda2e4)
O acúmulo de fluidos e de células da defesa no local de infecção causa vermelhidão, inchaço, aquecimento e dor, coletivamente denominados inflamação. A inflamação é, tradicionalmente, definida por quatro palavras do latim (calor, dolor, rubor e tumor) que significam calor, dor, rubor e inchaço (edema), o que reflete os efeitos das citocinas e de outros mediadores inflamatórios nos vasos sanguíneos locais. Os neutrófilos e os macrófagos são os principais tipos de células presentes na fase inicial de uma resposta inflamatória, sendo também conhecidos como células inflamatórias. Como os macrófagos, os neutrófilos possuem receptores de superfície para constituintes bacterianos comuns e para o complemento, e são as principais células que capturam e destroem os microrganismos invasores. O influxo de neutrófilos é logo seguido por monócitos, que rapidamente se diferenciam em macrófagos, reforçando e mantendo a resposta imune inata. Mais lentamente, os eosinófilos também migram para o tecido inflamado e também contribuem para a destruição do microrganismo invasor.
Os mastócitos também desempenham um papel importante na inflamação aguda, pois expressam uma grande variedade de receptores de reconhecimento padrão e contêm grandes grânulos pré-formados de mediadores inflamatórios. Ainda, a inflamação determina o acúmulo de fibrina, que forma um envoltório ao redor do local, evitando a progressão da infecção.
Algumas citocinas liberadas no local da inflamação, tais como a interleucina-1 (IL-1), alcançam o centro termorregulador localizado no hipotálamo, originando a febre (elevação da temperatura corporal). Apesar do mal-estar e desconforto, a febre é um importante fator no combate às infecções, pois além de ser desfavorável para a sobrevivência dos microrganismos invasores, também estimula muitos dos mecanismos de defesa de nosso corpo.
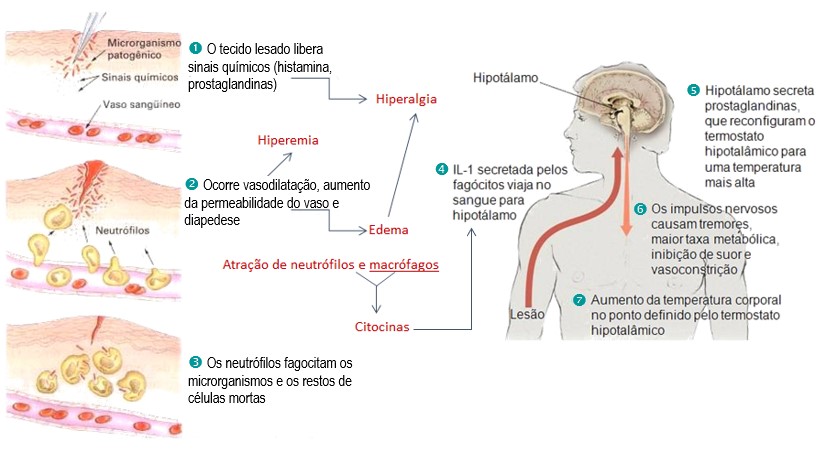
A inflamação de um tecido infectado tem vários efeitos benéficos no combate à infecção. Ela recruta células e moléculas da imunidade inata da circulação para os tecidos onde são necessárias para destruir o patógeno diretamente. Além disso, aumenta o fluxo de linfa contendo microrganismos e células portadoras de antígenos para os tecidos linfoides vizinhos, onde ativarão os linfócitos e iniciarão a resposta imune adaptativa.
Finalmente, uma vez que a resposta imune adaptativa foi ativada, a inflamação também recruta os efetores do sistema imune adaptativo, as moléculas de anticorpo e as células T efetoras para os locais de infecção.
Citocinas
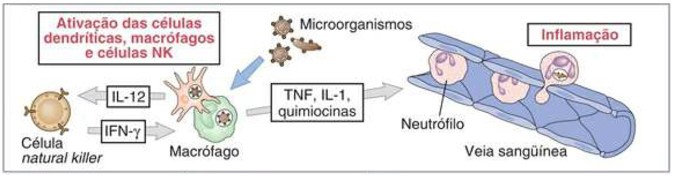
Em resposta aos patógenos, os macrófagos, as células dendríticas e outras células secretam citocinas, que são intermediárias em muitas reações celulares da imunidade inata.
Complementado o que foi mencionado previamente, as citocinas são proteínas solúveis que servem de mediadoras nas reações imunológicas e inflamatórias, sendo responsáveis pela comunicação entre leucócitos e entre os leucócitos e outras células. A maioria das citocinas é chamada, por convenção, de interleucina (IL). Apesar de o termo interleucina significar que essas moléculas são produzidas pelos leucócitos e atuam nos leucócitos, muitas citocinas são produzidas ou atuam em outras células, além dos leucócitos, e muitas citocinas que preenchem esses critérios têm outros nomes por motivos históricos.
Na imunidade inata, as fontes principais das citocinas são as células dendríticas e os macrófagos ativados ao reconhecerem os microrganismos. A ligação de componentes bacterianos (como o lipopolissacarídeo – LPS) ou moléculas virais (como o RNA de filamento duplo) a receptores das células dendríticas e dos macrófagos que reconhecem características comuns de muitos patógenos, é um estímulo poderoso para que essas células produzam citocinas que estimulam a inflamação (recrutamento dos leucócitos) e ativam as células natural killer (NK), as quais, por sua vez, produzem o interferon gama (IFN-), uma citocina que ativa os macrófagos e participa tanto da imunidade inata quanto da adquirida.
OBS.:
As citocinas também são produzidas na imunidade celular. Nesse tipo de imunidade adquirida, as principais fontes de citocinas são os linfócitos T auxiliares.
Na Tabela a seguir são enumeradas algumas características importantes das principais citocinas da imunidade inata. O nome fator de necrose tumoral (TNF) originou-se de uma experiência que demonstrou que uma citocina induzida pelo lipopolissacarídeo destruía tumores em ratos. Atualmente sabemos que esse efeito resulta da trombose dos vasos sanguíneos do tumor induzida pelo TNF, que é uma forma exagerada de uma reação que ocorre na inflamação. O nome interferon refere-se à capacidade de essas citocinas "interferirem" na infecção viral. O IFN- apresenta uma reação antiviral fraca quando comparado com os interferons (IFNs) do tipo I (INF- e INF-).
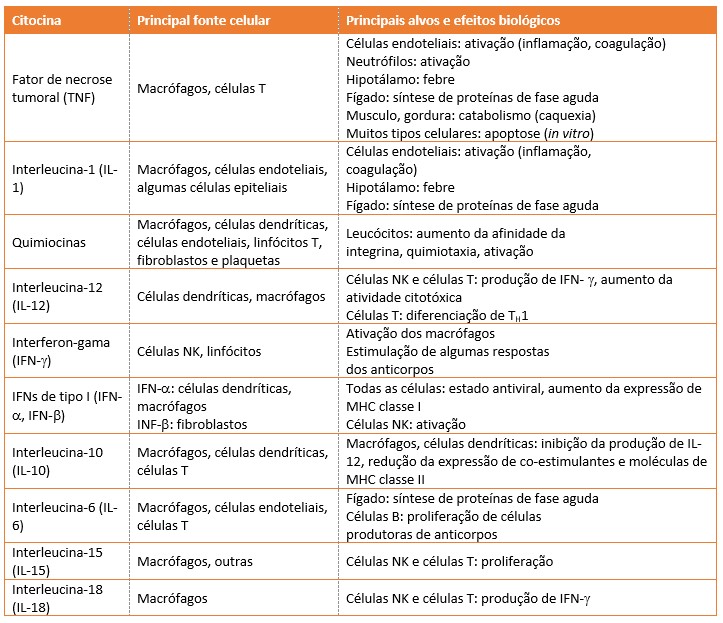
IFN= interferon; LPS= lipopolissacarídeo; MHC= complexo principal de histocompatibilidade; NK= natural killer.
Para maiores informações sobre citocinas, consulte:
- PALOMINO, D.C.T.; MARTI, L.C. Quimiocinas e imunidade. Einstein 2015; 13(3): 469-473.
O sistema do complemento (SC)
O sistema do complemento (SC) é um conjunto de proteínas circulantes e de membrana celular que desempenham papéis importantes na defesa do hospedeiro contra microrganismos e na lesão tecidual mediada por anticorpos. É constituído por uma família de mais de 20 glicoproteínas plasmáticas sintetizadas no fígado (principalmente), macrófagos e fibroblastos. Cada componente ativado do sistema adquire atividade proteolítica, ativando os elementos seguintes em cascata. Ao longo do processo, ocorre a produção de diversos mediadores que alteram a permeabilidade vascular e contribuem para o desenvolvimento da resposta inflamatória. Finalmente, ocorre formação do complexo de ataque à membrana (MAC, do inglês membrane attack complex), que promove a lise osmótica da célula-alvo, favorecendo a eliminação do agente infeccioso.
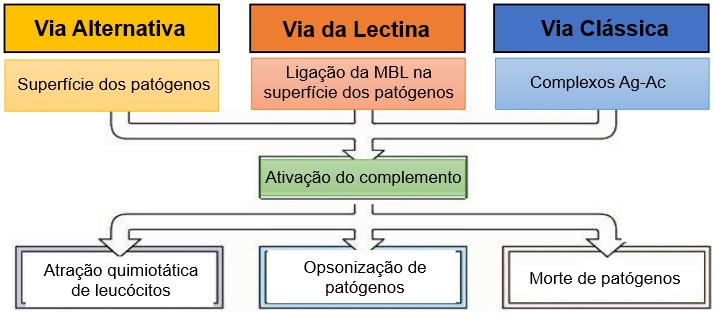
A cascata do complemento pode ser ativada por três vias: via alternativa, via da lectina ligante de manose (MBL, do inglês mannose binding lectin) e via clássica A ativação dessas vias contribui para a integração dos mecanismos efetores da imunidade inata e adaptativa. Na resposta imune inata, patógenos que invadem o organismo deparam com substâncias solúveis da resposta imune inata, como as proteínas do sistema do complemento, a proteína C reativa, entre outras. Na imunidade adaptativa, o SC é ativado pela ligação de anticorpos ao patógeno ou antígeno (imunocomplexo ou complexo antígeno-anticorpo ou complexo Ag-Ac).

OBS.:
Os componentes da via clássica costumam ser rotulados com um C e um número (p. ex., C1, C2, C3…), com base na ordem em que essas proteínas foram identificadas. Já na via alternativa, os componentes são geralmente classificados por letras (p. ex., fator B, fator D) ou por nome (p. ex., properdina).
Existem diversas proteínas de cada via que interagem numa sequência exata. As três vias de ativação do complemento diferem em como são iniciadas, mas compartilham as etapas finais, desempenhando as mesmas funções efetoras. A proteína do complemento mais abundante no plasma e que desempenha um papel central em todas as três vias é chamada de C3. O principal fragmento da clivagem proteolítica de C3 pelas enzimas geradas nas etapas iniciais, chamado de C3b, se liga de maneira covalente a microrganismos, sendo capaz de ativar proteínas do complemento presentes na superfície bacteriana.
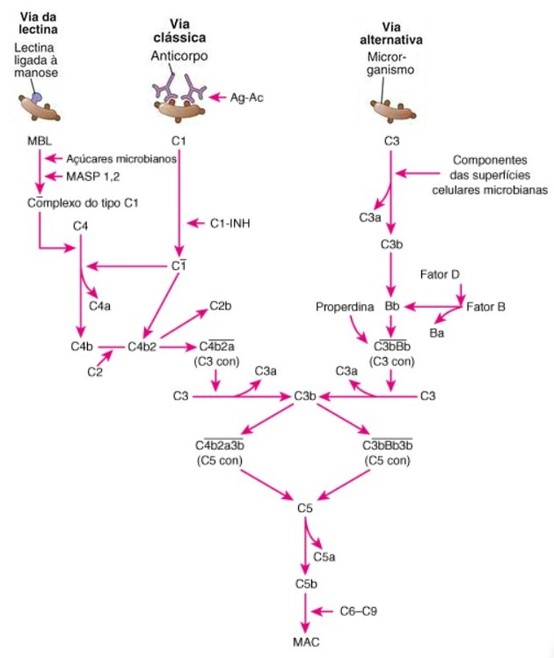
A via alternativa é um componente da imunidade inata, sendo desencadeada quando algumas proteínas do complemento são ativadas na superfície dos microrganismos. Se inicia com a quebra espontânea do componente C3 nos fragmentos C3a e C3b por componentes da superfície celular de microrganismos. A clivagem expõe uma ligação tioéster no fragmento C3b, que permite sua ligação covalente à superfície dos microrganismos invasores. A ligação de C3b permite a ligação ao fator B, que, em seguida, é clivado nos fragmentos Ba e Bb pelo fator D. O complexo C3bBb (C3 convertase da via alternativa) cliva mais moléculas C3 e permanece ligado na superfície. Esse complexo é estabilizado pela properdina (fator P), amplificando a quebra de C3. C3bBb cliva o componente C3, gerando C3bBbC3b, uma protease capaz de clivar C5.
A via da lectina também é um componente da imunidade inata, e é ativada quando uma proteína plasmática, a lectina ligante de manose (MBL), se liga à manose terminal nas glicoproteínas da superfície dos microrganismos. Essa via tem início pelo reconhecimento de manose na superfície de microrganismos pela MBL ligada às serina-proteases MASP1 e MASP2. A ativação dessas proteases resulta na quebra dos componentes C2 e C4 do sistema do complemento em fragmentos menores (C2b e C4a) e fragmentos maiores (C2a e C4b). O complexo C4bC2a constitui a C3 convertase (C3 con) da via clássica, que cliva C3 em C3a solúvel e C3b, que, por sua vez, se liga a C4bC2a na superfície do microrganismo. O complexo C4bC2aC3b, denominado C5 convertase (C5 con), cliva o componente C5, dando sequência a essa via, que culmina com a formação do MAC.
OBS.:
A MBL é estruturalmente similar a um componente de C1 da via clássica e serve para ativar C4. As etapas subsequentes são em essência as mesmas encontradas na via clássica. A lectina ativa proteínas da via clássica, mas como é iniciada por um produto bacteriano (na ausência de anticorpos), é um componente da imunidade inata.
A via clássica se inicia pela ligação do componente C1q a duas moléculas de IgG ou a uma de IgM, complexadas ao antígeno-alvo (ocorre quando C1 interage com o complexo Ag-IgM ou agrega os complexos Ag-IgG). Essa ligação ativa as proteases R (C1r) e S (C1s) associadas a C1q, que clivam os componentes C2 e C4. O complexo resultante, C4b2b, se liga de modo covalente ao anticorpo e à superfície microbiana onde o anticorpo está ligado. Ocorre, então, uma ruptura de C3, e o C3b gerado se liga de novo ao microrganismo. Parte de C3b se liga ao complexo C4b2b, e o complexo C4b2b3b funciona como C5 convertase, dando sequência à via, que culmina com a formação do MAC. Essa via é regulada pelo inibidor de C1 (C1-INH).
OBS.:
A via clássica está associada à resposta imune humoral adaptativa, pois depende da produção prévia de anticorpos específicos aderidos à superfície dos patógenos.
O resultado líquido dessas etapas iniciais da ativação do complemento é que os microrganismos adquirem um revestimento com ligação covalente a C3b. Devemos lembrar que a via alternativa e a via da lectina são mecanismos efetores da imunidade inata, e que a via clássica é um mecanismo da imunidade humoral adaptativa. Essas vias diferem no modo como elas são iniciadas, mas depois que são desencadeadas suas etapas posteriores são as mesmas.
As etapas posteriores da ativação do complemento são iniciadas pela ligação de C5 a C5 convertase, e pela proteólise de C5, que dá origem a C5b. Os componentes restantes, C6, C7, C8 e C9, se ligam na sequência. A proteína final na via, C9, se polimeriza para formar um poro na membrana celular, permitindo a passagem de água e íons, o que causa a morte da célula. Este poli-C9 é chamado de complexo de ataque de membrana (MAC), e sua formação é o resultado final da ativação do complemento.
O sistema do complemento tem três funções na defesa do hospedeiro. Em primeiro lugar, o C3b reveste os microrganismos, ligando-os às células fagocitárias por meio de receptores para o C3b expressos nos fagócitos. Assim, os microrganismos que são opsonizados (recobertos) com as proteínas do complemento são fagocitados rapidamente e destruídos pelos fagócitos. Em segundo lugar, alguns fragmentos proteolíticos das proteínas do complemento, em especial C5a e C3a, são quimioatrativos para fagócitos, promovendo o recrutamento dos leucócitos (inflamação) no local da ativação do complemento. Em terceiro lugar, a ativação do complemento culmina na formação de um complexo proteico polimérico que se insere na membrana celular microbiana, perturbando a permeabilidade da barreira que leva à lise osmótica ou à morte apoptótica do microrganismo.
Para maiores informações sobre imunidade inata, consulte:
- CRUVINEL, W.M.; MESQUITA JÚNIOR, D.; ARAÚJO, J.A.P.; CATELAN, T.T.T.; SOUZA, A.W.S.; SILVA, N.P.; ANDRADE, L.E.C. Sistema Imunitário – Parte I: Fundamentos da imunidade inata com ênfase nos mecanismos moleculares e celulares da resposta inflamatória. Rev Bras Reumatol 2010; 50(4): 434-461.