

Os sentidos somáticos
A sensibilidade somática origina-se da estimulação de receptores sensoriais localizados na pele, tela subcutânea, túnicas mucosas (da boca, vagina e ânus), músculos, tendões e articulações.
Os receptores sensoriais para a sensibilidade somática estão distribuídos irregularmente: algumas partes da superfície do corpo são densamente povoadas com receptores, enquanto outras contêm apenas alguns poucos.
A sensibilidade somática que se origina da estimulação da superfície da pele são as sensações cutâneas. As áreas com maior densidade de receptores sensitivos cutâneos são a ponta da língua, os lábios e as pontas dos dedos. Uma das funções mais importantes da pele, graças à sua grande extensão e à sua abundante inervação sensorial, é receber estímulos do meio ambiente.
A pele e as sensações cutâneas
A pele é o maior órgão do corpo humano, chegando a medir 2 m2 e pesar 4 Kg em um adulto. É constituída por duas camadas distintas, firmemente unidas entre si – a epiderme (mais externa, formada por tecido epitelial) e a derme (mais interna, formada por tecido conjuntivo).
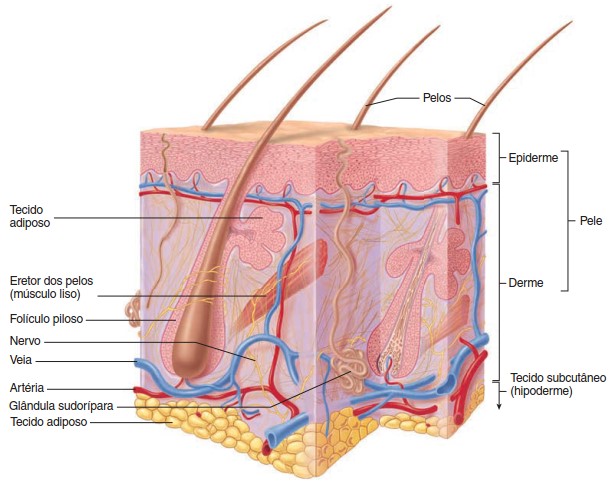
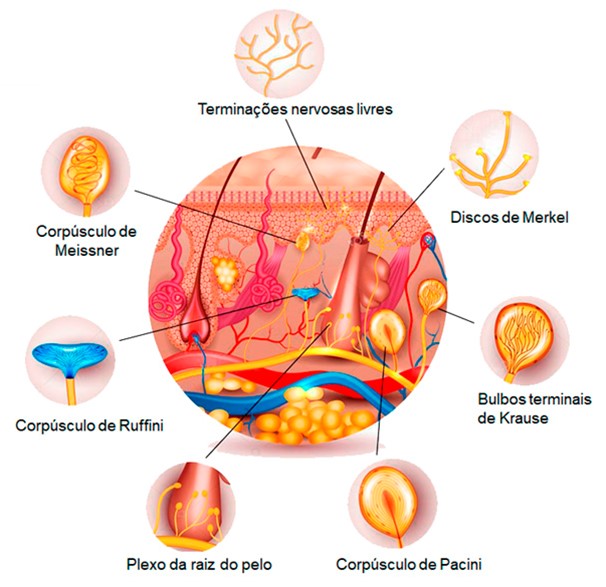
Uma vez que toda a superfície cutânea é provida de terminações nervosas capazes de captar estímulos térmicos, mecânicos ou dolorosos, a pele também é o maior órgão sensorial que possuímos, sendo suficientemente sensível para discriminar um ponto em relevo com apenas 0,006 mm de altura e 0,04 mm de largura quando tateado com a ponta do dedo. Essas terminações nervosas ou receptores cutâneos são especializados na recepção de estímulos específicos. Não obstante, alguns podem captar estímulos de natureza distinta. Cada receptor tem um axônio e, com exceção das terminações nervosas livres, todos eles estão associados a tecidos não neurais.
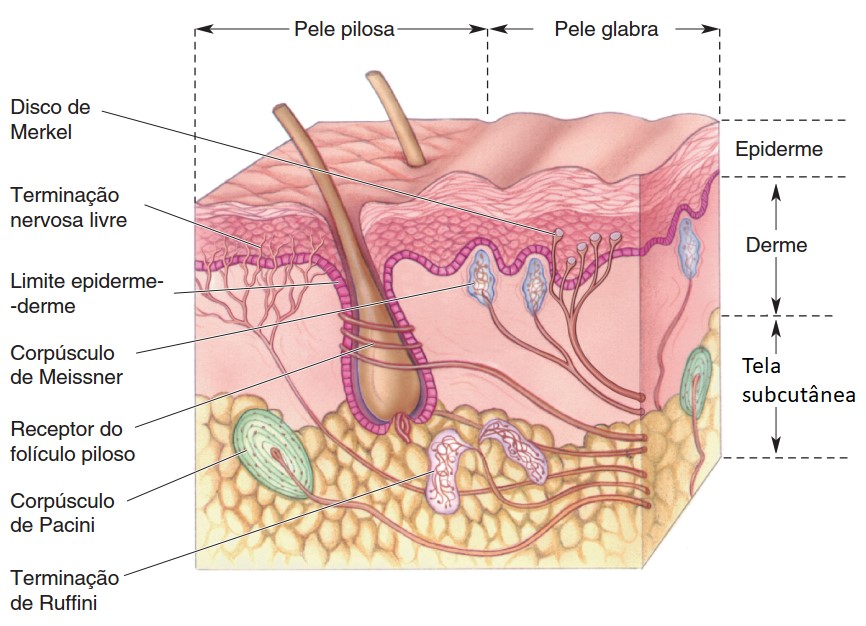
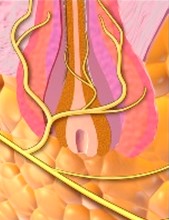
Nas regiões da pele providas de pelo, existem terminações nervosas específicas nos folículos capilares formadas por axônios que envolvem o folículo piloso (plexo da raiz pilosa), as quais captam as forças mecânicas aplicadas contra o pelo.
Na pele desprovida de pelo (pele glabra) e também na que está coberta por ele, encontram-se ainda três tipos de receptores comuns:
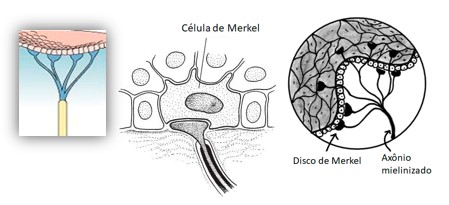
1) Células e discos de Merkel: sensibilidade tátil e de pressão. Uma fibra aferente costuma estar ramificada com vários discos terminais destas ramificações nervosas. Estes discos estão englobados em uma célula especializada, cuja superfície distal se fixa às células epidérmicas por um prolongamento de seu protoplasma. Assim, os movimentos de pressão e tração sobre epiderme desencadeiam o estímulo.
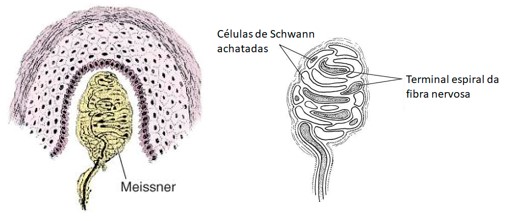
2) Corpúsculos de Meissner: tato, pressão, vibração (detectam vibrações de baixa frequência). Estão nas saliências da pele sem pelos (como nas partes mais altas das impressões digitais). São formados por um axônio mielínico, cujas ramificações terminais se entrelaçam com células acessórias.
3) Terminações nervosas livres: sensíveis aos estímulos mecânicos (cócegas, prurido ou coceira), térmicos (calor e frio) e especialmente aos dolorosos. São formadas por um axônio ramificado envolto por células de Schwann (mielinizado) sendo, por sua vez, ambos envolvidos por uma membrana basal.
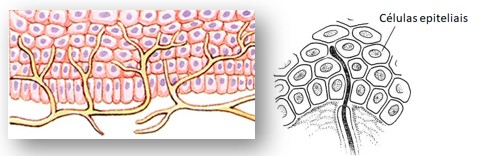
4) Corpúsculos de Pacini (ou Vater-Pacini): captam especialmente pressão e vibração (detectam vibrações de alta frequência). São formados por uma fibra nervosa cuja porção terminal, amielínica, é envolta por várias camadas que correspondem a diversas células de sustentação. A camada terminal é capaz de captar a aplicação de pressão, que é transmitida para as outras camadas e enviada aos centros nervosos correspondentes.
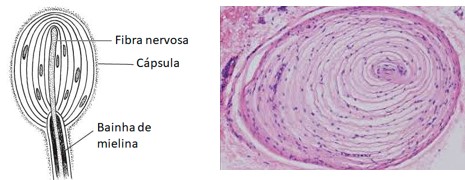
Na pele sem pelo (pele glabra) encontram-se ainda outros receptores específicos:
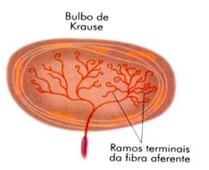
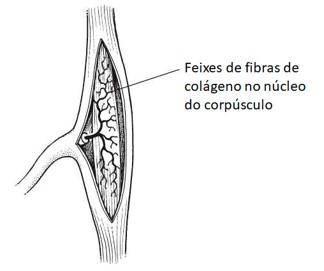
5) Bulbos terminais de Krause: receptores térmicos de frio. São formados por uma fibra nervosa cuja terminação possui forma de clava. Situam-se nas regiões limítrofes da pele com as membranas mucosas (por exemplo: ao redor dos lábios e dos genitais).
6) Corpúsculos de Ruffini: receptores táteis. São receptores encapsulados alongados e em forma de fuso localizados profundamente na derme, nos ligamentos e nos tendões. Embora estruturalmente semelhantes a outros receptores táteis, não são bem compreendidos. Eles representam cerca de 20% dos receptores na mão humana (pele glabra) e não provocam nenhuma sensação tátil específica quando estimulados eletricamente. Embora ainda haja dúvidas quanto à sua função, eles provavelmente respondem principalmente a estímulos gerados internamente.
| Receptor de superfície | Sensação percebida |
| Células e discos de Merkel | Tato e pressão |
| Receptores de Meissner | Tato, pressão, vibração (detectam vibrações de baixa frequência) |
| Terminações nervosas livres | Cócegas, prurido (coceira), calor, frio e dor |
| Plexos das raízes pilosas | Tato |
| Receptores de Pacini (ou Vater-Pacini) | Pressão, vibração (detectam vibrações de alta frequência) |
| Receptores de Krause (Bulbos terminais de Krause) | Frio |
| Receptores de Ruffini | Tato |
| Modalidade do estímulo | Estímulo | Tipo de receptor | Receptor sensorial |
| Tato | Tato e pressão | Mecanorreceptor | Receptores de Merkel, Meissner, Pacini e Ruffini |
| Temperatura | Quantidade de calor | Termorreceptor | Receptores de Krause (frio) e Terminações nervosas livres (frio e calor) |
| Dor | Estímulos intensos e substâncias químicas | Nociceptor | Terminações nervosas livres |
Veja também:
- O que provoca as cócegas? Por que a gente ri, mesmo sendo desconfortável?
- O tegumento como receptor sensorial.
Mediadores químicos da dor
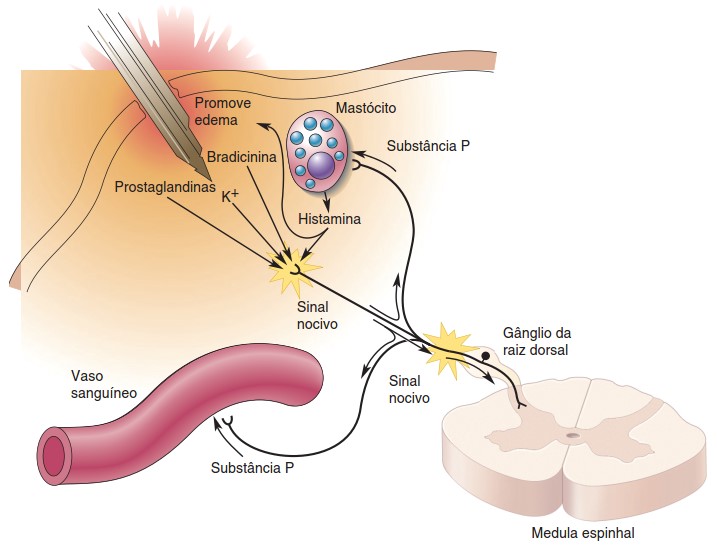
A sensação somática depende demais de nociceptores, que são as terminações nervosas livres ramificadas e não mielinizadas que sinalizam lesão ou risco de lesão ao corpo.
Vários produtos químicos modulam a excitabilidade dos nociceptores, tornando-os mais sensíveis aos estímulos térmicos ou mecânicos que provocam dor:
- bradicinina: despolariza diretamente os nociceptores e estimula mudanças celulares que deixam mais sensíveis os canais iônicos ativados pela temperatura;
- prostaglandinas: gerados pela quebra enzimática de lipídeos de membrana. Não desencadeiam diretamente a dor, mas aumentam muito a sensibilidade dos nociceptores a outros estímulos;
- substância P: peptídeo sintetizado pelos próprios nociceptores. Causa vasodilatação e liberação de histamina a partir dos mastócitos e também pode provocar a sensibilização de outros nociceptores ao redor do local da lesão.
A ativação de um ramo do axônio de um nociceptor pode levar à secreção de substância P por outros ramos daquele axônio nas vizinhanças. As informações sensoriais, após chegarem à medula espinhal, são transmitidas ao bulbo, tálamo e finalmente córtex somatossensorial.
Clique no link para assistir ao vídeo: Como seu cérebro responde à dor
A sensibilidade Proprioceptiva
A sensibilidade proprioceptiva permite que saibamos a localização da cabeça e dos membros e como estão se movimentando, mesmo quando não estamos olhando para eles, assim, podemos caminhar, digitar ou vestir sem usar os olhos. Cinestesia é a percepção dos movimentos do corpo. A sensibilidade proprioceptiva origina-se nos receptores denominados proprioceptores.
Os proprioceptores fornecem informações com relação à posição e ao movimento do corpo. Os proprioceptores embutidos nos músculos (especialmente nos músculos posturais) e nos tendões nos informam o grau de contração dos músculos, a quantidade de tensão nos tendões e as posições das articulações. O aparelho vestibular, na orelha interna, monitora a orientação da cabeça em relação ao solo e a posição da cabeça durante os movimentos. A forma pela qual fornecem informações para a manutenção do equilíbrio e da estabilidade.
Os proprioceptores também nos permitem fazer a discriminação com base no peso, a capacidade de avaliar o peso de um objeto. Esse tipo de informação ajuda a determinar o esforço muscular necessário para realizar uma tarefa. Por exemplo, se pegarmos um objeto, rapidamente percebemos o seu peso e, em seguida, exercemos a quantidade correta de esforço necessário para levantá-lo.
Como os proprioceptores se adaptam de forma lenta e apenas levemente, o encéfalo continuamente recebe impulsos nervosos relacionados com a posição de diferentes partes do corpo e faz os ajustes para assegurar a coordenação.
Os três tipos de proprioceptores são:
- fusos musculares: dentro dos músculos esqueléticos;
- órgãos tendíneos (corpúsculos tendíneos de Golgi ou órgãos tendinosos): dentro dos tendões;
- receptores cinestésicos articulares: dentro das cápsulas articulares.
Fusos musculares
Os fusos musculares são os proprioceptores presentes nos músculos esqueléticos que monitoram as mudanças no comprimento dos músculos esqueléticos e participam nos reflexos de estiramento.
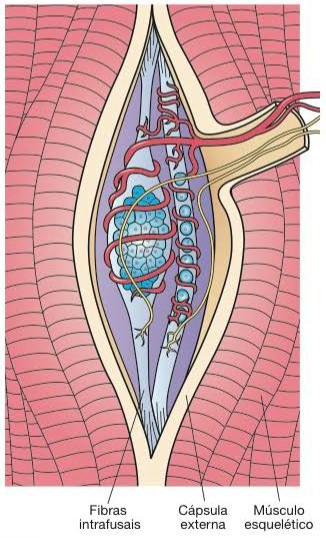
Eles são constituídos por uma cápsula de tecido conjuntivo que delimita um espaço contendo fluido e fibras musculares modificadas, denominadas fibras intrafusais. Diversas fibras nervosas sensoriais penetram os fusos musculares, onde detectam modificações no comprimento (distensão) das fibras musculares intrafusais e transmitem essa informação para a medula espinhal, onde são ativados diversos mecanismos reflexos que atuam sobre determinados grupos musculares. participando do mecanismo de controle da postura e da coordenação de músculos opostos durante as atividades motoras, como caminhar ou correr, por exemplo.
Um reflexo de estiramento opera da seguinte forma:
- O leve estiramento de um músculo estimula os fusos musculares, que monitoram as mudanças no comprimento do músculo.
- Em resposta ao estiramento, um fuso muscular gera um ou mais impulsos nervosos que se propagam ao longo de um neurônio sensorial somático pela raiz posterior do nervo espinhal e pela medula espinhal.
- Na medula espinhal (centro de integração), o neurônio sensorial faz uma sinapse excitatória com um neurônio motor, consequentemente, ativando-o no corno anterior da substância cinzenta.
- Se a excitação é forte o bastante, um ou mais impulsos nervosos originam-se no neurônio motor e se propagam ao longo de seu axônio, que se estende da medula espinal até a raiz anterior, e pelos nervos periféricos até o músculo estimulado. Os terminais axônicos do neurônio motor formam junções neuromusculares (JNM) com as fibras musculares esqueléticas do músculo estirado.
- A liberação de acetilcolina pelos impulsos nervosos nas JNM dispara um ou mais potenciais de ação muscular no músculo estirado (efetor) e o músculo se contrai. Assim, o estiramento muscular é acompanhado pela contração muscular, que anula o estiramento.
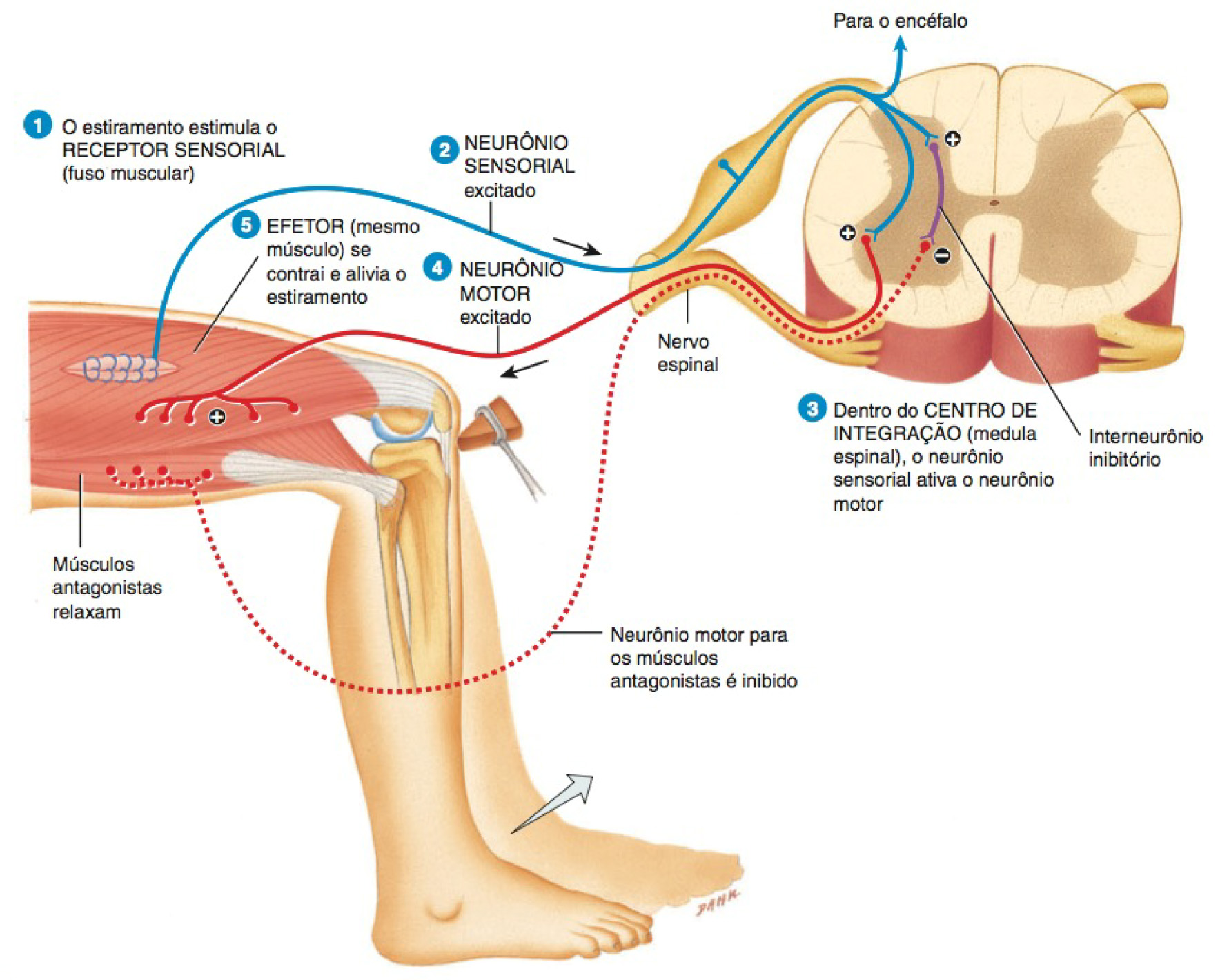
Ao ajustar a intensidade de resposta de um fuso muscular ao estiramento de um músculo esquelético, o encéfalo estabelece o nível geral do tônus muscular, o menor grau de contração presente enquanto o músculo está em repouso.
Corpúsculos tendíneos de Golgi
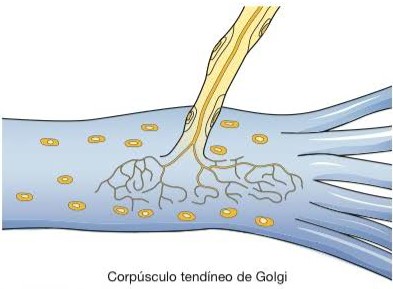
Nas junções de um tendão com um músculo, os tendões apresentam feixes de fibras colágenas encapsuladas, nas quais penetram fibras nervosas sensoriais, constituindo os órgãos tendíneos ou corpúsculos tendíneos de Golgi, que detectam e respondem às mudanças na tensão muscular, provocadas pelo estiramento passivo ou contração muscular.
Quando é aplicada tensão a um músculo, os órgãos tendíneos geram impulsos nervosos que se propagam pelo SNC, fornecendo informações com relação às alterações na tensão muscular. Os reflexos tendíneos diminuem a tensão muscular, produzindo relaxamento.
Ao iniciar os reflexos tendíneos, os órgãos tendíneos protegem os tendões e seus músculos associados contra a lesão decorrente de tensão excessiva (quando um músculo se contrai, exerce uma força que traciona os pontos de fixação, em ambas as extremidades, na direção um do outro; essa força é a tensão muscular).
Um reflexo tendíneo opera da seguinte forma:
- À medida que a tensão aplicada a um tendão aumenta, o órgão tendíneo (receptor sensorial) é estimulado (despolarizado até o limiar).
- Impulsos nervosos originam-se e propagam-se na medula espinhal ao longo de um neurônio sensitivo.
- Dentro da medula espinhal (centro de integração), o neurônio sensitivo ativa um interneurônio inibitório que faz sinapse com um neurônio motor.
- O neurotransmissor inibitório inibe (hiperpolariza) o neurônio motor que, em seguida, gera uns poucos impulsos nervosos.
- O músculo relaxa e alivia o excesso de tensão.
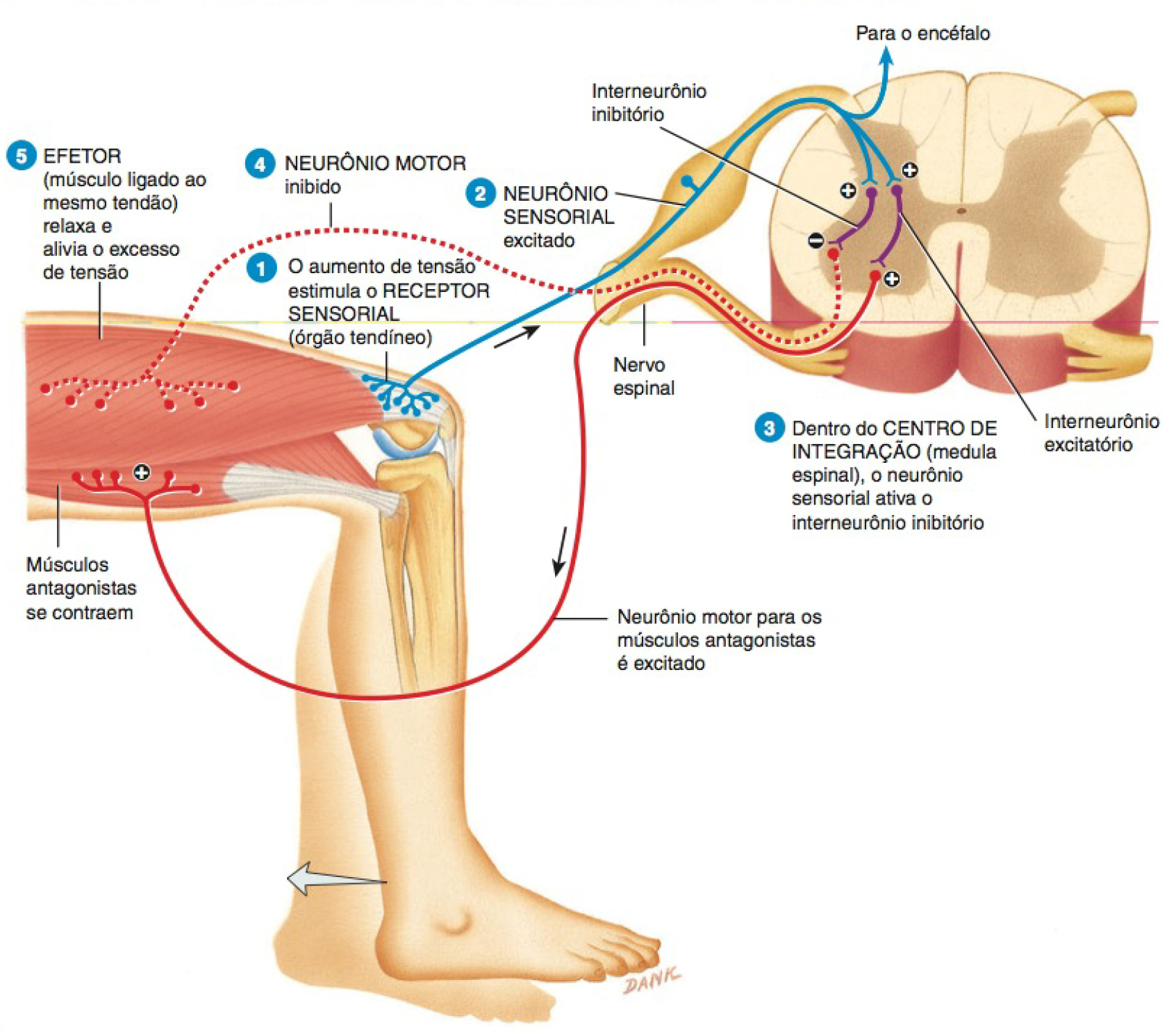
Portanto, à medida que a tensão no órgão tendíneo aumenta, a frequência de impulsos inibitórios aumenta; a inibição dos neurônios motores para o músculo que desenvolve excesso de tensão (efetor) provoca o relaxamento do músculo. Dessa forma, o reflexo tendíneo protege o tendão e o músculo dos danos decorrentes de tensão excessiva.
Receptores cinestésicos articulares
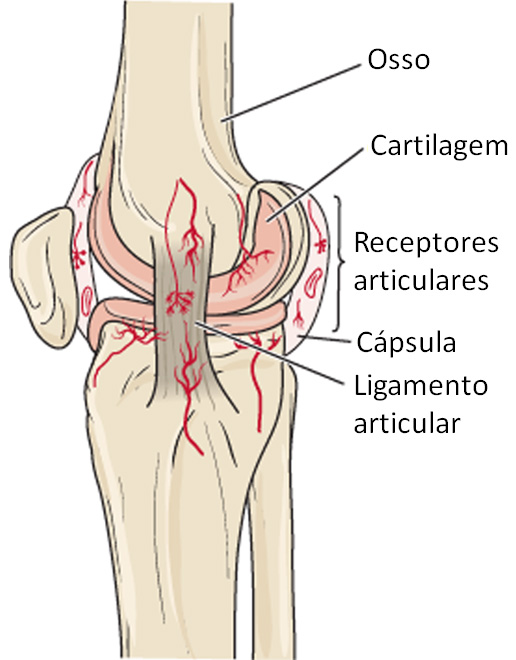
Diversos tipos de receptores cinestésicos articulares estão presentes no interior e em torno das cápsulas articulares das articulações sinoviais. Terminações nervosas livres e mecanorreceptores cutâneos tipo II nas cápsulas articulares respondem à pressão.
Os corpúsculos lamelados no tecido conjuntivo fora das cápsulas articulares respondem à aceleração e à desaceleração durante o movimento. Os ligamentos articulares contêm receptores semelhantes aos órgãos tendíneos, que ajustam a inibição reflexa dos músculos adjacentes quando é exercida tensão excessiva sobre a articulação.