

Componentes e mecanismos da imunidade adquirida (adaptativa)
A resposta imune inata inicialmente defende o corpo contra infecções, mas atua somente para controlar os patógenos que possuam certas características moleculares ou que induzam a liberação de determinadas citocinas e outras moléculas de defesa não-específicas secretadas. Desta forma, algumas infecções podem ser combatidas apenas pela imunidade inata, e os patógenos serão eliminados precocemente, produzindo alguns sintomas e causando pouco dano. No entanto, a maioria dos patógenos pode sobrepujar o sistema imune inato e, nesse caso, a imunidade adaptativa é essencial para combatê-los, sendo iniciada quando os receptores antigênicos dos linfócitos reconhecem os antígenos.
Os linfócitos B e T diferem em relação aos antígenos que reconhecem. As moléculas de reconhecimento de antígenos dos linfócitos B são os anticorpos ou imunoglobulinas (Ig) ligados à membrana das células B. Estas proteínas atuam como receptores celulares para antígenos e são chamadas de receptores de células B (BCRs). Podem reconhecer uma grande variedade de macromoléculas (proteínas, polissacarídeos, lipídios e ácidos nucléicos), assim como pequenas substâncias químicas em solução ou associadas à superfície celular, sendo que cada linfócito B produz uma imunoglobulina de especificidade única. Por outro lado, as moléculas de reconhecimento de antígenos dos linfócitos T (receptores de células T ou TCRs) podem identificar somente fragmentos peptídicos de antígenos proteicos, e assim mesmo apenas quando estes são apresentados por moléculas especializadas nas células do hospedeiro. Consequentemente, as respostas imunológicas mediadas pelas células T só podem ser geradas contra os antígenos proteicos dos microrganismos que estão associados às células do hospedeiro.
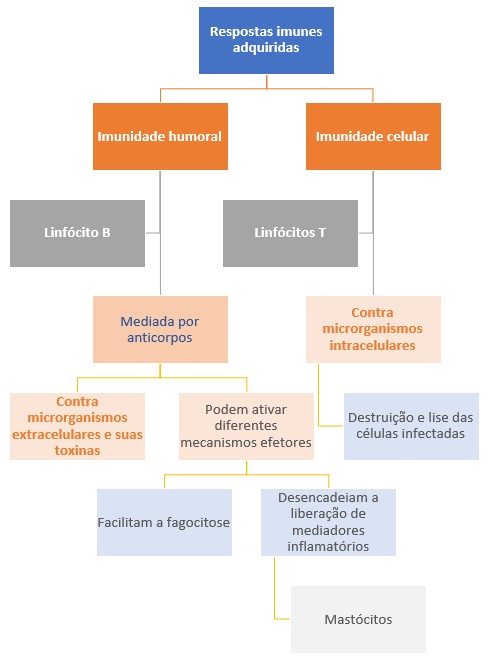
Portanto, existem dois tipos de imunidade adquirida, conhecidos como imunidade humoral e imunidade celular, que são mediados por moléculas e células diferentes e projetados para fornecer a defesa contra microrganismos extracelulares e intracelulares, respectivamente.
A imunidade humoral é mediada pelos anticorpos (ou imunoglobulinas) produzidos pelos linfócitos B diferenciados, os plasmócitos. Os anticorpos são secretados na circulação e nos líquidos das mucosas, neutralizando e eliminando os microrganismos e as toxinas microbianas presentes fora da célula do hospedeiro, no sangue e no lúmen dos órgãos mucosos, como os tratos gastrointestinal e respiratório. Uma das funções mais importantes dos anticorpos é impedir que patógenos presentes nas mucosas e no sangue tenham acesso e colonizem as células e os tecidos conjuntivos do hospedeiro, evitando, assim, que as infecções se estabeleçam. Porém, os anticorpos não têm acesso aos microrganismos que vivem e se multiplicam dentro de células infectadas.
A defesa contra os microrganismos intracelulares é chamada de imunidade celular porque é mediada pelas células conhecidas como linfócitos T. Alguns linfócitos T ativam os fagócitos para destruir os microrganismos ingeridos pelas células fagocitárias nas vesículas fagocíticas. Outros linfócitos T destroem qualquer tipo de célula do hospedeiro que apresente microrganismos infecciosos em seu citoplasma.
Assim, os anticorpos produzidos pelos linfócitos B reconhecem os antígenos microbianos extracelulares, enquanto os linfócitos T reconhecem os antígenos produzidos pelos microrganismos intracelulares.
Na imunidade humoral, os linfócitos B secretam anticorpos que eliminam os microrganismos extracelulares. Na imunidade mediada por células, os linfócitos T ativam os macrófagos para matar os microrganismos fagocitados ou destroem as células já infectadas.
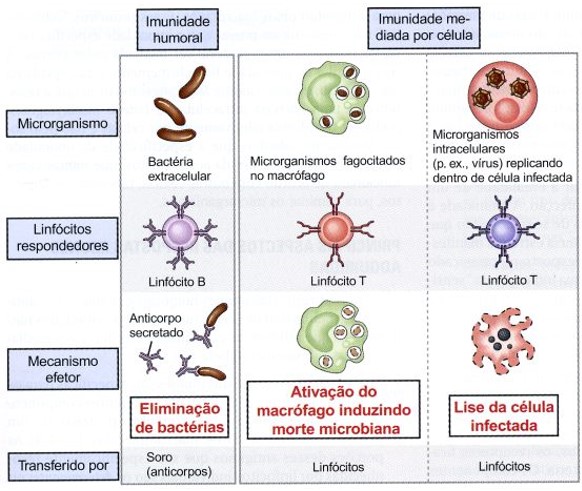
O objetivo da resposta adaptativa é ativar esses mecanismos de defesa contra os microrganismos que estão em localizações anatômicas diferentes, tais como a luz intestinal, a circulação ou dentro das células. Todas as respostas imunológicas adaptativas se desenvolvem em etapas, cada qual correspondendo a uma reação particular dos linfócitos.
Linfócitos T
Os linfócitos T são divididos em subpopulações funcionalmente distintas que reconhecem e respondem apenas a antígenos peptídicos associados à superfície da célula – os linfócitos T auxiliares (T helper – CD4+ ou T4), os linfócitos T supressores e os linfócitos T citotóxicos (CTLs ou citolíticos – CD8+).
As células T auxiliares, em resposta à estimulação antigênica, secretam citocinas, cuja função é a de estimular a proliferação e a diferenciação das células T e de outros tipos celulares, incluindo células B e macrófagos. Os CTLs lisam as células que produzem antígenos estranhos, como é o caso de certas células tumorais e de células infectadas por vírus e outros microrganismos intracelulares.
Os linfócitos T auxiliares estimulam a transformação dos linfócitos B em plasmócitos. Os linfócitos T supressores inibem as respostas humoral e celular e apressam o término da resposta imunitária. Linfócitos T auxiliares e T supressores são células reguladoras. Os linfócitos T citotóxicos agem diretamente sobre as células estranhas e as infectadas por vírus, graças a dois mecanismos. Um deles é a produção de proteínas chamadas perforinas, que abrem orifícios nas membranas plasmáticas, provocando a lise das células. Pelo outro mecanismo, os linfócitos T citotóxicos induzem as células-alvo a entrarem no processo de morte celular programada, chamado apoptose.
O centro dessas funções é a especificidade restrita dos linfócitos T para os antígenos que são exibidos pelas células infectadas do hospedeiro. No timo, os linfócitos T se diferenciam nas subpopulações das células T auxiliar, T supressora e T citotóxica. Em outros locais se podem formar as células T da memória.
Células apresentadoras de antígenos (APCs)
.jpg?alt=media&token=22a6ecc2-8bd3-406d-84d5-234f466a0ff1)
Como visto anteriormente, os linfócitos T podem identificar somente fragmentos peptídicos de antígenos proteicos que estão associados às células do hospedeiro, e, mesmo assim, apenas quando estes são apresentados por proteínas especializadas, que são codificadas pelo complexo principal de histocompatibilidade (MHC – do inglês major histocompatibility complex). As moléculas do MHC humanas são chamadas antígenos leucocitários humanos (HLA – do inglês human leucocyte antigens), porque foram primeiramente descobertas nos leucócitos do sangue.
Células apresentadoras de antígenos (APCs, do inglês antigen-presenting cells) são células do sistema de defesa inato, como os macrófagos e monócitos, as células dendríticas, e as células epiteliais do timo, capazes de fagocitar ou internalizar um antígeno, processá-lo e expô-lo em sua superfície para que sejam apresentados aos linfócitos T (os neutrófilos, assim como os macrófagos e as células dendríticas, são representantes do grupo de células fagocitárias do sistema imune, mas, diferentemente destas células, os neutrófilos não apresentam antígenos para os linfócitos T).
Um aspecto característico das APCs é a expressão de moléculas do MHC (HLA). Os linfócitos B, embora não sejam parte do sistema imune inato, também funcionam como APCs, pois têm receptores antígeno-específicos que permitem que elas internalizem, processem e apresentem grandes quantidades de antígeno específicos.
O processamento do antígeno é essencial para a ativação dos linfócitos T, pois essas células não reconhecem as moléculas antigênicas nativas (não processadas). As células T só reconhecem antígenos quando associados a moléculas MHC, enquanto as células B reconhecem as moléculas antigênicas (proteínas, pequenos peptídeos, lipídios, polissacarídeos e moléculas menores) diretamente, sem necessidade de qualquer tratamento prévio.
O MHC está dividido em três classes – I, II e III – e ocupa uma região de aproximadamente 4 megabases (Mpb) no braço curto do cromossomo 6 (6p21.3). Seus mais de 220 genes são herdados em blocos ou séries chamados haplótipos e expressos codominantemente em cada indivíduo. Desta forma, os MHC têm uma estrutura que é única para cada pessoa, e esse é o principal motivo pelo qual enxertos e transplantes de órgãos são rejeitados, exceto quando feitos entre gêmeos univitelinos (gêmeos idênticos), que têm constituição molecular e MHC idênticos.
As regiões de classes I e II do MHC codificam dois grupos de proteínas estruturalmente distintas, porém homólogas. As moléculas do MHC de classe I apresentam os peptídeos às células T CD8+ (linfócitos T citotóxicos ou citolíticos) e as moléculas de classe II apresentam os peptídeos às células T CD4+ (linfócitos T auxiliares). Uma vez estimuladas, as células T CD4+ produzem citocinas capazes de estimular outros linfócitos T e outras importantes células de defesa.
Para maiores informações sobre MHC, consulte:
- MIRANDA-VILELA, A.L. A diversidade genética do complexo principal de histocompatibilidade (MHC) e sua relação com a susceptibilidade para doenças autoimunes e câncer. Ribeirão Preto/SP, Sociedade Brasileira de Genética (SBG), 2007.
Anticorpos e imunidade humoral
Em princípio, quase todas as estruturas químicas podem ser reconhecidas pelo sistema imune adaptativo como um antígeno, mas os antígenos geralmente encontrados em uma infecção são proteínas, glicoproteínas e polissacarídeos de patógenos. Um receptor antigênico individual ou um anticorpo reconhece uma pequena parte da estrutura molecular da molécula antigênica, que é conhecida como determinante antigênico ou epítopo.
Como discutido anteriormente, anticorpos são as formas secretadas dos receptores de antígenos das células B. São glicoproteínas plasmáticas circulantes do tipo das gamaglobulinas, que, por isso, são também chamadas de imunoglobulinas (Ig). Visto que são solúveis e secretados em grandes quantidades, os anticorpos são facilmente obtidos e estudados. Por essa razão, a maior parte do que sabemos sobre os receptores das células B vem do estudo de anticorpos.
Cada anticorpo interage especificamente com o determinante antigênico (epítopo) que estimulou sua formação. Os anticorpos são secretados pelos plasmócitos que surgem pela proliferação e diferenciação de linfócitos B.
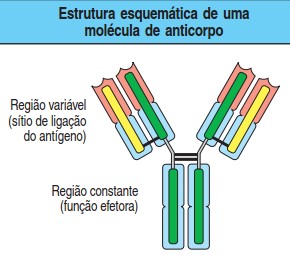
Cada molécula de anticorpo, é composta de quatro cadeias polipeptídicas, sendo duas cadeias pesadas (H) idênticas (representadas em verde na figura) e duas cadeias leves (L) idênticas (representadas em amarelo na figura). Na região variável, cada cadeia leve está ligada a uma cadeia pesada, e as duas cadeias pesadas estão ligadas uma à outra por pontes dissulfeto. Entre essas regiões existem as dobras ou dobradiças, regiões que permitem o dobramento das cadeias polipeptídicas, dando flexibilidade à molécula de imunoglobulina.
Assim, as moléculas de anticorpos possuem uma forma de Y e consistem em três segmentos de igual tamanho, conectados por uma porção flexível. As duas extremidades dos dois braços do Y formam dois sítios idênticos de ligação antigênica, mas são variáveis entre diferentes moléculas de anticorpo – a região V, ao passo que a base do Y, ou região C, é mais conservada e interage com as moléculas e as células efetoras. Tanto as cadeias leves como as pesadas apresentam, em suas regiões variáveis, as chamadas regiões hipervariáveis, correspondendo a zonas de intensa diversidade.
Se a molécula da imunoglobulina for clivada enzimaticamente (com papaína ou pepsina, p. ex.), resultam vários fragmentos. Dois deles são semelhantes, resultando da clivagem com papaína; cada um contém um sítio capaz de se combinar com o antígeno específico e é denominado de fragmento que se combina com o antígeno ou Fab, (do inglês, antigen binding fragment). O terceiro fragmento não tem poder combinatório com o antígeno e pode ser cristalizado, sendo por isso chamado de Fc (do inglês, crystallizable fragment) ou fragmento constante; liga-se ao complemento e a receptores de muitos tipos diferentes de células envolvidas na resposta imune, sendo responsável por muitas atividades biológicas que ocorrem após a ligação ao antígeno.
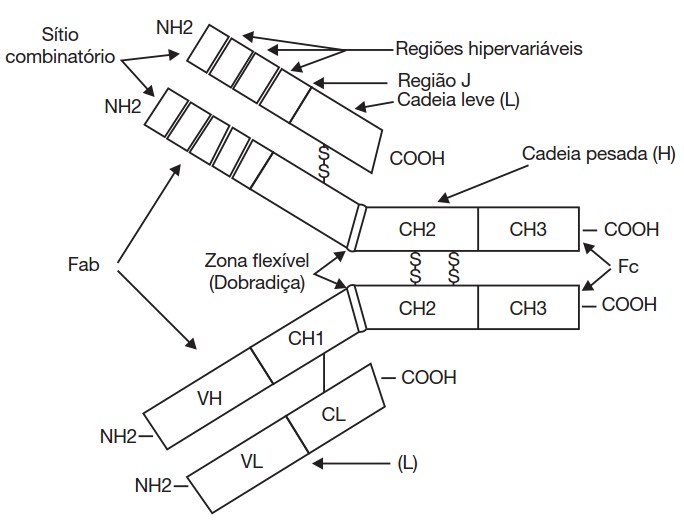
Portanto, as moléculas de anticorpos são compostas por duas regiões distintas. Uma é a região variável que pode ser composta por uma variedade infinita de diferentes sequências de aminoácidos, formando estruturas sutilmente diferentes que permitem que o anticorpo se ligue especificamente a uma ampla variedade de antígenos. A outra é a região constante, que apresenta apenas uma de cinco formas bioquimicamente distinguidas, e estas são conhecidas como classes ou isotipos de anticorpos. Na espécie humana, as cinco classes principais de imunoglobulinas são IgA, IgD, IgE IgG e IgM.
Características das imunoglobulinas | |||
| Classe | % no sangue | Presente em | Funções conhecidas |
| IgA | 10-16 | Secreções (lágrimas, saliva, muco), sangue e linfa | Proteção das superfícies mucosas |
| IgD | 0,2 | Superfície dos linfócitos B, sangue e linfa | Receptor de antígenos em células B virgens (BCR); co-expressa com IgM na superfície desses linfócitos |
| IgE | 0,002 | Sangue | Participa das reações alérgicas; atua contra protozoários parasitas |
| IgG | 80 | Sangue, linfa e intestino | Facilita a fagocitose, neutraliza toxinas, protege o feto (único anticorpo que atravessa a placenta) |
| IgM | 5-10 | Superfície dos linfócitos B, sangue e linfa | Ativo contra microrganismos; aglutina antígenos; o primeiro a ser produzido na resposta imune |
A região constante determina as propriedades funcionais de um anticorpo – como ele irá participar dos mecanismos que eliminam os antígenos uma vez que ele é reconhecido – e cada classe realiza sua função particular participando de um grupo distinto de mecanismos efetores.
A imunoglobulina mais abundante no plasma é a lgG, que constitui cerca de 75% das imunoglobulinas do plasma sanguíneo. IgG é a única imunoglobulina que atravessa a barreira placentária humana e passa para o sangue fetal, contribuindo para a defesa imunitária do recém-nascido.
IgM constitui 10% das imunoglobulinas do plasma sanguíneo e é a imunoglobulina que predomina no início das respostas imunitárias. Junto com a IgD (que constitui apenas 0,2% do total de imunoglobulinas), a IgM é a principal imunoglobulina encontrada na superfície dos linfócitos B, onde funcionam como receptores que se combinam com antígenos específicos. Essa combinação resulta na proliferação dos linfócitos B e sua diferenciação em plasmócitos. A IgM circulante pode ativar o complemento, culminando na lise de bactérias, além de outros efeitos.
A IgA existe em pequena quantidade no plasma sanguíneo, mas sob a forma de IgA secretora (SlgA), é o principal anticorpo encontrado nas secreções como lágrima, leite, saliva, secreções nasal e brônquica, secreção contida no lúmen do intestino delgado, secreção da próstata e, também, no líquido que lubrifica a vagina. A SlgA é muito resistente às enzimas proteolíticas e, por isso, adaptada para atuar nas secreções sem sofrer inativação pelas enzimas lá encontradas.
A IgE tem grande afinidade para receptores localizados na membrana dos mastócitos e basófilos. Imediatamente após sua secreção pelos plasmócitos, as moléculas de IgE prendem-se àqueles receptores e, praticamente, desaparecem do plasma. A reação alérgica é mediada pela atividade da IgE e dos antígenos (alergênios) que estimulam sua produção. Quando o alergênio que provocou a produção de IgE é novamente encontrado, o complexo antígeno-IgE que se forma na superfície dos mastócitos e basófilos determina a produção e a liberação de diversas substâncias biologicamente ativas, como histamina, heparina, leucotrienos e ECF-A (eosinophil chemotactic factor of anaphylaxis).
Veja também:
- Edema de glote é reação alérgica grave que afeta a respiração e pode matar.
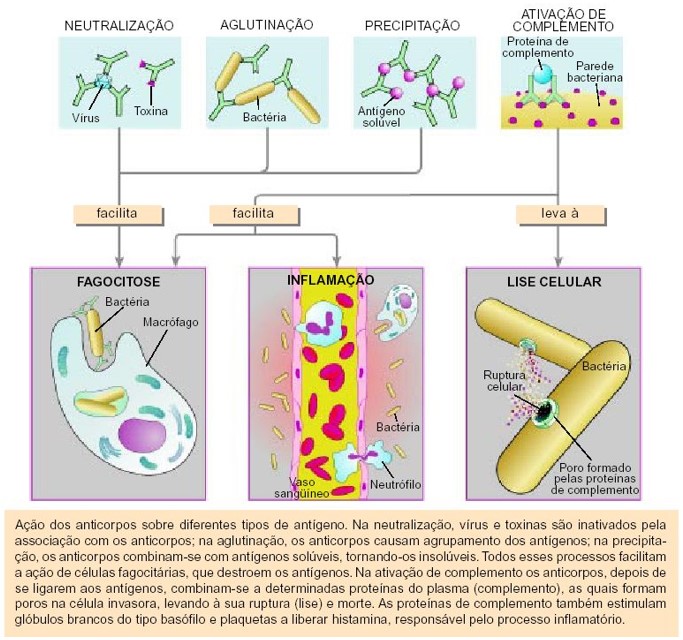
Os anticorpos apresentam diversos mecanismos de ação, dos quais destacaremos os mais importantes. Uma importante função do anticorpo é se combinar especificamente com o epítopo que ele reconhece e, então, provocar o aparecimento de sinais químicos, indicando a presença do invasor aos outros componentes do sistema imunitário. Alguns anticorpos podem aglutinar células e precipitar antígenos solúveis. A aglutinação de microrganismos facilita sua fagocitose, e a precipitação de moléculas agressivas estranhas (p. ex., toxinas) pode torná-las inócuas. Os antígenos que se ligam aos anticorpos IgG e IgM ativam o complemento, um grupo de proteínas do plasma sanguíneo que causam a lise (ruptura da membrana) dos microrganismos. Uma vez ativado, o complemento também facilita a fagocitose de bactérias e outros microrganismos patógenos (causadores de doenças). As bactérias ligadas ao complexo antígeno-IgG são ditas opsonizadas. A opsonização facilita muito a fagocitose.
Para maiores informações sobre imunidade adquirida, consulte:
- MESQUITA JÚNIOR, D.; ARAÚJO, J.A.P.; CATELAN, T.T.T.; SOUZA, A.W.S.; CRUVINEL, W.M.; ANDRADE, L.E.C.; SILVA, N.P. Sistema Imunitário – Parte II: Fundamentos da resposta imunológica mediada por linfócitos T e B. Rev Bras Reumatol 2010; 50(5):552-580.